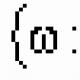Peroxidação lipídica da membrana neuronal. Peroxidação lipídica (LPO)
MINISTÉRIO DA EDUCAÇÃO DA REPÚBLICA DA BIELORRÚSSIA
UNIVERSIDADE MÉDICA DO ESTADO DE GOMEL
Faculdade de Medicina e Diagnóstico
Departamento de Diagnóstico Laboratorial Clínico
Diagnóstico laboratorial da intensidade da peroxidação lipídica
Trabalho do curso
Intérprete: Podstrekha Elena Stanislavovna
aluno do grupo D-503
Supervisor científico: chefe do departamento, Doutor em Ciências Médicas, Professora Associada Novikova Irina Aleksandrovna
GOMEL 2016
Ensaio
O trabalho analisa a literatura sobre o estudo das vias enzimáticas e não enzimáticas de formação de espécies reativas de oxigênio, os mecanismos de seus efeitos prejudiciais às células vivas, em particular o início da peroxidação lipídica por radicais livres. São considerados sistemas antioxidantes e pró-oxidantes de defesa do organismo, equilibrando a formação, o metabolismo e a utilização de espécies reativas de oxigênio.
Curso 35 páginas, 1 tabela, 17 fontes.
Lista de palavras-chave: peroxidação lipídica, oxidação de radicais livres, espécies reativas de oxigênio, proteção antioxidante, malondialdeído.
Lista de abreviações
Introdução
O papel da oxidação dos radicais livres e do sistema antioxidante no corpo humano
1 Formas de radicais livres no corpo
2 Características gerais das principais ERO, seu papel biológico
3 Papel fisiológico da oxidação dos radicais livres
4 Regulação da oxidação de radicais livres
5 O sistema de defesa antioxidante do corpo
6 Mecanismos patogenéticos de distúrbios que se desenvolvem quando o equilíbrio dos sistemas antioxidante e pró-oxidante é perturbado
Radicais livres (peroxidação lipídica)
1 Participação de EROs e produtos de peroxidação lipídica na patogênese de doenças humanas
2 Diagnóstico de processos de peroxidação
Conclusão
Lista de símbolos
AOA – atividade antioxidante
AO - antioxidante
ROS - espécies reativas de oxigênio
GR - glutationa redutase
GP - glutationa peroxidase
MP - mieloperoxidase
LPO - peroxidação lipídica
SOD - superóxido dismutase
FRO - oxidação radical livre
CL - quimiluminescência
EPR - ressonância paramagnética eletrônica
Introdução
peroxidação lipídica oxigênio ativo
No organismo, em decorrência de reações redox, ocorre constantemente a geração de espécies reativas de oxigênio (ROS), que são altamente reativas, causando, principalmente, a modificação oxidativa de biopolímeros: proteínas, lipídios, ácidos nucléicos, carboidratos. Os radicais de oxigênio, apesar de sua reatividade e potencial toxicidade, em baixas concentrações são metabólitos normais de muitas reações bioquímicas na célula. Sob condições fisiológicas, as reações dos radicais livres ocorrem em níveis baixos. Os processos que ocorrem com a participação dos radicais de oxigênio indicam o importante papel desses compostos na manutenção da homeostase, na formação da resistência do organismo às infecções e na garantia da regeneração de tecidos e órgãos. Se o processo de geração de ROS se intensificar, isso pode ser e é um fator desencadeante para o desenvolvimento de toda uma lista de vários processos patológicos.
A relevância do desenvolvimento aprofundado do problema do significado patogenético da peroxidação dos radicais livres é determinada pelo aumento do sofrimento ambiental. O estudo deste importante elo da homeostase tem significado prático direto, pois permite desenvolver e aplicar abordagens preventivas adequadas que previnam o início de uma reação em cadeia de oxidação dos radicais livres ou neutralizem a toxicidade dos produtos da peroxidação lipídica.
1. O papel da oxidação dos radicais livres e do sistema antioxidante no corpo humano
Os sistemas envolvidos na formação de ERO e os processos associados à alteração oxidativa de compostos biológicos são convencionalmente unidos pelo conceito de sistema pró-oxidante.
Os pró-oxidantes em uma célula viva incluem altas concentrações de oxigênio (por exemplo, durante a oxigenação hiperbárica prolongada de um paciente), sistemas enzimáticos que geram radicais superóxido (por exemplo, xantina oxidase, enzimas da membrana plasmática dos fagócitos, etc.), íons ferrosos.
As reações oxidativas são a base da produção de energia e da atividade vital de todas as células do corpo humano. Podem ocorrer sem adição de oxigênio (reações de oxidase) e com adição de oxigênio molecular ou atômico - reações de oxigenase. Os produtos intermediários destes últimos são peróxidos e epóxidos, portanto tais reações são chamadas de peroxidação. É induzido por radicais livres altamente reativos.
Em condições fisiológicas, a intensidade dos processos peróxidos é insignificante e mantida em um nível estacionário graças a um sistema multicomponente para neutralizar os radicais livres formados constantemente - o sistema antioxidante.
A formação de pró-oxidantes nos tecidos é equilibrada pela atividade de antioxidantes intra e extracelulares, formando um certo nível ideal de equilíbrio pró-oxidante-antioxidante.
1.1 Formas de radicais livres no corpo
Existem muitas reações redox conhecidas que produzem vários tipos de radicais livres. Os radicais livres foram descobertos no final do século passado. Até o momento, mais de 8.000 deles foram descritos. Os radicais livres são compostos altamente ativos que se formam em condições fisiológicas como produtos secundários durante o metabolismo, bem como de outras formas, incluindo reações redox realizadas por transferência de um único elétron; homólise de moléculas iniciadoras com ligações covalentes fracas, radiólise; fotólise, termólise.
Muitos xenobióticos, incluindo várias drogas, álcool, etc., são metabolizados no corpo, gerando radicais livres. Apesar da grande diversidade de origem e estrutura, os radicais livres são divididos em 3 grandes grupos: 1) radicais reativos de oxigênio (ROS); 2) radicais reativos de nitrogênio (RNS); 3) radicais reativos de cloro (RCS).
Um radical livre é uma molécula, átomo ou grupo de átomos que possui um elétron desemparelhado em seu orbital atômico externo. As ROS são participantes ativos em um grande número de reações químicas nas células, exercendo uma variedade de efeitos fisiológicos.
Características distintivas dos radicais livres:
a presença de um elétron desemparelhado no nível de energia externo;
próprio momento magnético;
alta atividade química e vida útil curta;
a capacidade de iniciar reações em cadeia de oxidação;
A ocorrência mais provável de radicais livres no corpo é durante a adição sequencial de elétrons ao oxigênio e durante a peroxidação lipídica dos radicais livres.
Os principais processos que levam à formação de radicais livres no corpo:
adição sequencial de elétrons ao oxigênio na presença de metais de valência variável;
oxidação microssomal e mitocondrial, fagocitose;
reações enzimáticas envolvendo hidrolases, oxidases, desidrogenases;
reações de autooxidação e biossíntese (tióis, catecolaminas, etc.);
oxidação de compostos estranhos - xenibióticos, alguns medicamentos;
a influência de fatores ambientais negativos (iniciadores de oxidação física e química);
processos fotoquímicos;
peroxidação lipídica.
modificação de propriedades físico-químicas de membranas biológicas;
funções protetoras, oxidação de compostos estranhos, efeito microbicida;
metabolismo, acumulação e biotransformação de energia;
influência na imunidade, transferência de informações.
As formas mais comuns de radicais livres no corpo são:
Espécies que reagem ao oxigênio:
SOBRE ˙ 2 - radical ânion superóxido; ¹ O2 - forma singlete de oxigênio; OH ˙ - radical hidroxila; H2O2 - peróxido de hidrogênio; Halogênios oxidados: CLO ˙ - hipocloreto, cloraminas; Óxidos de nitrogênio: NÃO - Óxido nítrico; Radicais livres formados durante a peroxidação lipídica: RO ˙ ,RO2 ˙ - radicais mono-, diméricos, poliméricos, cíclicos, alcoxi e peróxidos de ácidos graxos. 2 Características gerais das principais ERO, seu papel biológico Os radicais de oxigênio são formados durante processos associados ao transporte de elétrons ao longo da cadeia respiratória. Em condições normais, de 1 a 5% do oxigênio consumido é consumido para a geração de ERO. No entanto, esse valor pode aumentar significativamente quando o orçamento de oxigênio do corpo muda - durante hiperóxia ou hipóxia. Como resultado da redução sequencial do oxigênio molecular, ocorre a formação de ânion superóxido, peróxido de hidrogênio e radical hidroxila. Radical ânion superóxido (O ˙ 2) é formado quando um elétron é adicionado a uma molécula de oxigênio no estado fundamental. O radical é um agente oxidante relativamente fraco e em muitos sistemas biológicos atua como doador de elétrons, reduzindo vários compostos. Ao interagir com um próton, o O2 se transforma em um radical hidroperóxido. Além disso, é uma fonte potencial de radical hidroxila e peróxido de hidrogênio. O radical ânion superóxido é um composto mais reativo que o oxigênio. No corpo, o radical ânion superóxido é um produto intermediário de muitas reações bioquímicas, como a oxidação de tióis, flavinas, quinonas, catecolaminas, pterinas, bem como o metabolismo de xenobióticos. Das fontes do radical ânion superóxido, as mais interessantes são a hemoglobina, a mioglobina, o citocromo C reduzido, as NADPH oxidases das células fagocíticas, etc. A principal fonte do radical no sangue são os neutrófilos, que o geram durante uma série de reações específicas. e imunidade inespecífica. Outra enzima especializada na formação do radical ânion superóxido é a xantina oxidase, que em condições normais está predominantemente na forma desidrogenase e pode se transformar de forma reversível e irreversível na forma oxidase, que ocorre durante a isquemia. Acredita-se que a geração do radical pela xantina oxidase seja necessária para o metabolismo do ferro, regulação do tônus vascular e proliferação celular e fornecimento do potencial microbicida dos neutrófilos. O radical de superoxidação é o gatilho de uma cascata de reações de radicais livres que levam ao surgimento da maioria das EROs e produtos de peroxidação lipídica. Está envolvido na síntese de peptídeos quimiotáticos, aumenta a proliferação de linfócitos estimulada por mitógenos, inibe a ação do fator relaxante endotelial, pode danificar as membranas dos eritrócitos, inibir a síntese de Ca2+-ATPase, RNA e proteínas das células endoteliais, oxidar as proteínas séricas, enquanto em ao mesmo tempo, sua citotoxicidade direta é baixa. Para regular os níveis de O ˙ 2 nas células existe uma enzima antioxidante altamente específica - a superóxido dismutase, que tem a capacidade de acelerar significativamente a reação de dismutação do radical em peróxido de hidrogênio. Peróxido de hidrogênio (H2O2). A adição de dois elétrons a uma molécula de oxigênio ou de um elétron a um ânion O2 é acompanhada pela formação de um ânion O22ˉ duplamente carregado, que se transforma em HO2 ˙ ou peróxido de hidrogênio. O peróxido de hidrogênio é classificado como um agente oxidante de força média; na ausência de antioxidantes enzimáticos e íons metálicos de valência variável, é relativamente estável e pode migrar para células e tecidos. O peróxido de hidrogênio tem um efeito prejudicial limitado, causando, em particular, perturbação da homeostase do cálcio na célula. No corpo, suas fontes são reações envolvendo oxidases (xantina oxidase, L-aminoácido oxidase e várias outras), transferindo dois elétrons para uma molécula de oxigênio, bem como a reação de dismutação do radical ânion superóxido, catalisada pela superóxido dismutase . Esta reação é a fonte de quase 80% do peróxido de hidrogênio no local da inflamação. O peróxido de hidrogênio é um produto intermediário na formação da maioria das ERO. H2O2 é uma das fontes dos ROS mais tóxicos - radical hidroxila. Na presença da mieloperoxidase, formam-se hipohaletos altamente reativos a partir dela - HOC1, HOBr, HOJ, HOSCN. Em condições normais, as células dos mamíferos são bastante resistentes aos efeitos do peróxido de hidrogênio devido à presença de enzimas - glutationa peroxidase e catalase. As moléculas de H2O2 desempenham uma série de funções reguladoras na célula. O peróxido pode servir como um sinal metabólico para induzir a expressão de genes que sintetizam proteínas estruturais e funcionais na célula. Radical hidroxila (HO ˙) é a mais reativa e, consequentemente, tóxica de todas as ERO formadas em sistemas biológicos. O radical pode quebrar qualquer ligação de hidrocarboneto, e a taxa de sua interação com substratos orgânicos atinge valores iguais à taxa de difusão (ou seja, 107-1010 mol/s, que é 106 vezes maior do que para o radical ânion superóxido e peróxido de hidrogênio) . Comparado com outro AFK, MAS ˙ tem o maior potencial redox, igual a + 2,7, o que lhe permite atacar e destruir quaisquer macromoléculas quase no local no momento de seu aparecimento. A principal fonte de radical hidroxila é a reação de Fenton com a participação de metais de valência variável, principalmente com Fe2+, conforme esquema H2O2 + Fe+2 → Fe+3 + OH + OH ˙. A formação do radical também ocorre durante a oxidação do ácido araquidônico, na reação de Haber-Weiss, Osipov, durante a oxidação microssomal, nas reações com enzimas flavinas e CoQ. A redução reversa de Fe3+ é possível em reação com O2, bem como em interação com ácido ascórbico, glutationa, cisteína e outros compostos oxidantes. Foi demonstrado que os efeitos citotóxicos e carcinogênicos da radiação ionizante estão diretamente relacionados à formação de radicais hidroxila durante a radiólise da água. MAS ˙ também participa da implementação dos efeitos microbicidas e citotóxicos de granulócitos, monócitos e linfócitos T. Os radicais hidroxila causam danos aos ácidos nucléicos, proteínas, bem como a outras estruturas celulares, e inibem uma série de frações do complemento. Eles induzem a formação de radicais orgânicos e, assim, desencadeiam processos de peroxidação lipídica. Devido à alta inespecificidade das reações do radical com diversas moléculas orgânicas, sua interação é em cadeia. É importante notar que o corpo não possui sistemas enzimáticos especializados que tenham a capacidade de inativar os radicais hidroxila. Compostos de baixo peso molecular, como uracila, ácido úrico, salicilatos, glicose, dimetilsulfóxido, têm a capacidade de inibir OH ˙ radical apenas em concentrações suficientemente altas. Assim, em uma série de condições patológicas acompanhadas pela formação excessiva de ROS e, consequentemente, de radicais hidroxila, o organismo fica praticamente indefeso contra os efeitos nocivos deste composto. A prevenção de danos às estruturas celulares é realizada apenas através da redução da concentração de radicais - precursores OH, em particular, radical ânion superóxido e peróxido de hidrogênio. SOD e catalase têm a capacidade de destruir esses radicais precursores. Oxigênio singleto ( ¹O 2). É formado quando o spin de um dos elétrons do orbital p na molécula de oxigênio muda. Emergência ¹O 2 como subproduto foi observado em muitas reações enzimáticas envolvendo SOD, catalase e peroxidases, bem como em reações envolvendo a maioria das ERO. Assim, na reação de decomposição do peróxido de hidrogênio pela catalase a 1%, o oxigênio resultante aparece no estado singleto. O oxigênio singlete é altamente reativo e entra facilmente em reações oxidativas com compostos orgânicos. Freqüentemente atua como um indutor de reações de LPO. Embora ¹ O2 tem efeito citotóxico e participa da ação microbiocida dos granulócitos, sua contribuição nesses processos não é decisiva. Um dos inibidores mais eficazes do oxigênio singleto na célula é o beta-caroteno, cuja molécula pode extinguir cerca de 1.000 de suas moléculas antes de sofrer destruição oxidativa. As ROS atuam como mensageiros secundários nos processos vitais das células. Por estarem envolvidas na transdução de sinal, as ERO afetam componentes-chave dos processos metabólicos: fosforilação, metabolismo do Ca2+, modulação de fatores de transcrição, hidrólise de fosfolipídios. Durante quaisquer reações de estresse do corpo, acompanhadas por um estado de estresse oxidativo, as ERO estão envolvidas na transmissão de sinais de mensageiros primários para desencadear uma cascata de reações necessárias para adaptação e sobrevivência em condições extremas. Cada tecido possui uma certa capacidade tampão de AOP. Depende do estado da AOP do fluido intercelular e da própria célula e de seus componentes individuais. Alguns tecidos, pelas características de sua atividade funcional e metabólica, são altamente sensíveis ao estado de estresse oxidativo, isso se deve ao alto poder potencial do sistema pró-oxidante e à baixa capacidade tampão do AOP; Esses tecidos incluem o cérebro, a retina e os pulmões. Isso se deve à importante função reguladora que as ERO geradas e os metabólitos radicais desempenham nesses tecidos. No tecido cerebral, isso está associado à transmissão de sinais de excitação, à ocorrência de um potencial de ação e à ativação de sinapses. AFK - mensageiros secundários. A base metabólica de qualquer célula depende da natureza da informação proveniente do ambiente. Os portadores desta informação são mensageiros primários: hormônios, citocinas, neurotransmissores. Este processo é realizado através de sinalização celular ou transdução de sinal. E mensageiros secundários estão incluídos na transmissão do sinal através da membrana celular. As ROS participam ativamente como intermediários secundários. Eles desempenham um papel regulador no crescimento celular, apoptose, adesão celular, coagulação sanguínea, etc. Concentrações baixas (micromolares) de ROS aumentam o crescimento ou melhoram a resposta à estimulação do crescimento em muitos tipos de células, e os antioxidantes suprimem a proliferação celular normal. Baixas concentrações de H2O2 estimulam o crescimento de fibroblastos. A inibição da SOD ou glutationa peroxidase aumenta a proliferação celular. ELE ˙ , é um fator que aumenta a proliferação celular e a atividade da proteína quinase ativada por mitógeno (MAP quinase). Em concentrações fisiológicas, as ERO no papel de segundos mensageiros são formadas indiretamente através da interação ligante-receptor. Hormônios (insulina, angiotensina, hormônio da paratireóide, vitamina D), citocinas e fatores de crescimento podem atuar como tais ligantes. A formação de complexos ligante-receptor é acompanhada pela formação de EROs, que estão ativamente envolvidas na transdução de sinal, afetando componentes-chave dos processos metabólicos na célula. Os mensageiros primários regulam o nível de ERO na célula, ativando os processos de sua geração, por um lado, e reduzindo a atividade de unidades AOP individuais, por outro. As citocinas participam ativamente desse processo. As citocinas estimulam a liberação de ERO de muitos tipos de células, incluindo fibroblastos humanos, células epiteliais e endoteliais. ROS está associado à transmissão de sinal do fator de crescimento derivado de plaquetas, fator de crescimento epidérmico, fator de crescimento transformador β -1, fator de necrose tumoral. A participação da interleucina-1 e do interferon na transdução de sinal está associada à formação de O2ˉ, e do TNF - com H2O2. As ROS também atuam como mensageiros secundários nas células ósseas. TNF, interleucina-1, hormônio da paratireóide e vitamina D estimulam a formação de ERO devido à presença em osteoclastos NADPH oxidase. O peptídeo vasoativo (angiotensina II) exerce seu efeito nos processos de contração muscular e crescimento celular da musculatura lisa vascular por meio da geração de oxigênio intracelular ˙ 2ˉ. Fonte O ˙ 2ˉ estavam as NADH e NADPH oxidases, uma vez que ambas as enzimas são ativadas pela angiotensina. As ERO, como mensageiros secundários, estão envolvidas na regulação do metabolismo do Ca2+, na estimulação da fosforilação de proteínas e na ativação de fatores de transcrição. Na presença de oxidantes, o transporte de Ca2+ através dos canais de cálcio aumenta e a bomba de Ca2+ dependente de ATP é inibida. Oxidantes, aumentando a atividade de vários as proteínas quinases estão envolvidas na regulação de vários processos celulares, como mitogênese, adesão celular, apoptose, etc. Também envolvida na ação dos oxidantes como mensageiros secundários está a fosfolipase A2. Sua ativação por oxidantes envolve o envolvimento de diversas vias de transdução de sinal. O ácido araquidônico, como produto da fosfolipase A2, é um importante mediador de processos como inflamação, processos imunológicos, atividade da NADPH oxidase e coagulação sanguínea. Assim, existem 3 formas possíveis de ação dos oxidantes como mensageiros secundários nos processos da célula associados à formação de moléculas sinalizadoras: Influência na estrutura das membranas celulares. Influência no estado do depósito de Ca2+, que é acompanhado por sua mobilização do depósito e entrada no citosol. Ativação da fosfolipase A2. 1.3 Papel fisiológico da oxidação dos radicais livres Qualquer radical é um indutor de reações de radicais livres. Os processos de oxidação de radicais livres (FRO) com a participação de ROS em intensidade bastante baixa são processos metabólicos normais. Os radicais de oxigênio induzem processos de peroxidação lipídica necessários para os processos de renovação de fosfolipídios e regulação da permeabilidade da membrana celular. Uma importante função fisiológica das ERO é a ativação de uma série de proteínas de membrana e imunoglobulinas, bem como enzimas que regulam a mudança de vias metabólicas e a síntese de compostos de alta energia na célula. A fosforilação oxidativa e a taxa de divisão celular estão diretamente relacionadas aos processos FRO. O peróxido de hidrogênio pode atuar como um sinal metabólico para processos intracelulares que levam à oxidação de grupos SH específicos de proteínas quinases. Uma vez ativadas, essas proteínas translocam-se para o núcleo e induzem a expressão de grupos de genes, cujos produtos de expressão são responsáveis por diversas formas de reações de defesa celular. Além disso, o peróxido tem um efeito semelhante ao da insulina. Os produtos das reações dos radicais livres e LPO estão envolvidos na biossíntese de progesterona, hormônios esteróides e tireoidianos, leucotrienos, tromboxano A2, protrombina. Uma propriedade importante dos metabólitos ativos do oxigênio e, em particular, do radical ânion superóxido, é a regulação do metabolismo do tecido conjuntivo. As ERO estimulam a proliferação de fibroblastos, a síntese e degradação de colágeno e triptofano e estão envolvidas no metabolismo do ferro. Algumas das oxigenases, nomeadamente um grupo de enzimas denominado citocromo P-450 (actualmente existem mais de uma centena de isoformas), além da hidroxilação de compostos endógenos, utilizam oxigénio e uma série de ERO para a desintoxicação de xenobióticos lipofílicos. As monooxigenases dependentes do citocromo P-450 e o sistema de transporte de elétrons associado a elas introduzem ERO diretamente na molécula do substrato, o que leva à formação de um produto oxidado e mais hidrofílico. No corpo, as monooxigenases dependentes do citocromo P-450 desempenham uma série de funções importantes. Em primeiro lugar, esta é a biotransformação oxidativa (biossíntese ou degradação) de moléculas endobióticas lipofílicas endógenas (esteróides, retinóides, metabólitos do ácido araquidônico) e, em segundo lugar, a biotransformação de compostos químicos xenobióticos vindos de fora, que não participam de processos bioquímicos normais na célula e estão sujeitos a remoção. A principal reação da monooxigenase é sempre acompanhada pela formação de um radical ânion superóxido, peróxido de hidrogênio e, às vezes, de metabólitos ativos do substrato oxidado. Estas enzimas estão predominantemente presentes no retículo endoplasmático das células; o sistema enzimático é expresso ao máximo nos hepatócitos, glândulas supra-renais e gônadas. As ROS estão envolvidas na regulação do tônus vascular, inibindo o óxido nítrico endógeno. Uma das formas radicais de oxigênio - peroxinitrila, que é facilmente convertida nos produtos iniciais - óxido de nitrogênio e radical ânion superóxido, é uma forma de transporte para NO ˙, como resultado, o alcance de ação dessa molécula sinalizadora pode aumentar significativamente. A natureza radical livre do peroxinitrila é a razão para o enfraquecimento e nivelamento de numerosos efeitos fisiológicos do óxido nítrico. A interação do óxido nitroso e do radical ânion superóxido é acompanhada por um aumento acentuado no potencial oxidativo deste último. Os metabólitos do oxigênio participam das reações da imunidade celular e humoral como reguladores e efetores desses processos. Em particular, os radicais de oxigénio gerados por oxidases de granulócitos neutrófilos e fagócitos mononucleares desempenham um papel importante nos efeitos microbicidas, citotóxicos e imunorreguladores destas células. Neste caso, os principais efetores da ação microbicida são o radical hidroxila, o peróxido de hidrogênio e os hipohaletos. ROS estimulam a proliferação de células imunocompetentes. Foi comprovada a participação direta do radical ânion superóxido na formação de fatores quimiotáticos que causam ativação e migração de leucócitos para o local da inflamação. O radical hidroxila também tem a capacidade de induzir a síntese de peptídeos quimiotáticos, o que ao mesmo tempo aumenta a proliferação estimulada por mitógenos e subsequente diferenciação de linfócitos. Muitos processos importantes, como a geração de produtos finais do metabolismo das purinas e a degradação da dopamina, são acompanhados pela produção de ERO. A estimulação adrenérgica leva fisiologicamente a um aumento, e a estimulação colinérgica - a um enfraquecimento da produção de ERO endógenas, altera de forma oposta o potencial redox da célula e cria condições para um efeito permissivo, quando o mesmo sinal provoca uma resposta diferente das células, dependendo em seu estado redox. Por exemplo, o TNF causa morte ou proliferação celular, porque o fator de transcrição dependente dele é desencadeado apenas quando o potencial oxidativo das células-alvo muda. Realizando reações protetoras, as células (macrófagos e histiócitos) podem aumentar repetidamente a produção de EROs. Durante a fagocitose, ocorre uma “explosão metabólica” nos fagócitos, ou seja, aumento múltiplo no consumo de energia pela célula fagocítica. Uma parte significativa dessa energia é gasta pelas oxidases dependentes de NADPH na formação do radical superóxido. As ROS exercem um efeito bactericida nos fagolisossomos, porque Ao contrário das hidrolases lisossomais, as ERO são capazes de destruir paredes celulares bacterianas intactas e membranas celulares intactas. O mecanismo dependente de oxigênio do estágio final da fagocitose é mais importante que o hidrolítico; As ERO também são secretadas externamente durante o processo de exacitose, contando com sua capacidade de destruir o agente causador da alteração primária, por peroxidação das membranas das células vizinhas, realizar autodanos secundários e promover a produção de mediadores inflamatórios eicosanóides. 1.4 Regulação da oxidação de radicais livres A taxa de FRO e o conteúdo de radicais livres no corpo são normalmente mantidos em um determinado nível por um sistema de regulação complexo e em vários estágios. Ele pode distinguir condicionalmente fatores específicos e inespecíficos, cujo significado e contribuição mudam em diferentes estágios de oxidação. Fatores inespecíficos: mecanismos que regulam a quantidade e qualidade do substrato de oxidação e a sua disponibilidade; fatores que afetam os iniciadores de oxidação, em particular o estado dos metais de valência variável; propriedades físico-químicas de membranas biológicas; mecanismos que mantêm baixo teor de O2 nos tecidos. Mecanismos específicos: enzimas responsáveis pela formação e metabolismo de ROS (SOD, catalase, etc.); sistemas que utilizam produtos peróxidos (glutationa peroxidase, glutationa redutase, etc.); interceptadores de ROS (metionina, histamina, etc.); bioantioxidantes (tocoferol, ubiquinona, ceruloplasmina). 1.5 O sistema de defesa antioxidante do corpo Com base nas propriedades dos derivados radicais do oxigênio molecular e dos peróxidos orgânicos, proteger o corpo de seus efeitos nocivos é uma das tarefas mais importantes na manutenção da homeostase. O sistema para proteger tecidos e células de metabólitos tóxicos de oxigênio e produtos de peroxidação lipídica pode ser dividido em fisiológico (mecanismos que regulam o fornecimento e fornecimento de oxigênio às células) e bioquímico (o próprio sistema antioxidante do corpo, ou seja, uma ampla classe de compostos químicos que reduzem a atividade de processos de agentes oxidativos radicais). A componente fisiológica do sistema de defesa AO do organismo garante um equilíbrio entre a intensidade do transporte de oxigénio para as células e os processos metabólicos para a sua utilização benéfica e segura. Esses mecanismos para limitar as reações dos radicais livres são fornecidos por: a presença de uma cascata de níveis de pressão parcial de oxigênio, diminuindo dos alvéolos para as células de 100-105 para 8-10 mmHg. Art., isto é, 10-13 vezes. reduzindo a tensão de oxigênio em algumas estruturas subcelulares em 100-1000 vezes em comparação com a pressão parcial de oxigênio nos capilares. O processo é mediado por uma distância intercapilar relativamente grande e alta afinidade da citocromo oxidase pelo oxigênio; redução da microcirculação nos tecidos com aumento da pressão parcial de oxigênio no sangue arterial. O chamado “vasospasmo hiperóxico” que ocorre tem diversas causas. É dada grande importância à remoção do efeito vasodilatador do CO, bem como à redução da atividade das NO sintases e, consequentemente, à redução da produção do principal fator de vasodilatação - o óxido nítrico. A possibilidade de um efeito vasoconstritor direto do oxigênio também é reconhecida. O sistema bioquímico AO do corpo pode ser dividido em específico e inespecífico. Um sistema AO específico visa destruir ROS e os produtos de suas transformações posteriores. A ação do sistema AO inespecífico está associada à prevenção de condições e possibilidades de vazamento de elétrons e geração de ROS durante reações redox (como parte da fosforilação oxidativa) ou no processo de autooxidação de substratos (oxidação microssomal). Os efeitos patogênicos da LPO são resistidos por sistemas enzimáticos especializados e por vários compostos não enzimáticos. As enzimas AO específicas incluem superóxido dismutase, catalase, peroxidases e transferases dependentes de glutationa. Este grupo de enzimas, localizadas principalmente intracelularmente, tem a capacidade de destruir radicais livres e também participar na decomposição de hidroperóxidos de forma não radical. As enzimas de defesa antirradicais são caracterizadas por alta seletividade de ação dirigida contra determinados radicais; especificidade de localização celular e de órgãos, bem como o uso de metais como estabilizadores, que incluem cobre, zinco, manganês, ferro e vários outros. OxidantesAntioxidantesEspécies reativas de oxigênio: O ˙ 2 - radical ânion superóxido; ¹ O2 - forma singlete de oxigênio; OH ˙ - radical hidroxila; H2O2 - peróxido de hidrogênio; Halogênios oxidados: CLO ˙ - hipocloreto, cloraminas; Óxidos de nitrogênio: NÃO - Óxido nítrico; Radicais livres formados durante a peroxidação lipídica: RO ˙ ,RO2 ˙ - radicais mono-, diméricos, poliméricos, cíclicos, alcoxil e peróxido de ácidos graxos [.Enzimáticos Superóxido dismutases Catalase Glutationa (GSH-)-peroxidases GSSG redutases Glutationa-S-transferases UDP-glucuroniltransferases NADP-N-quinona oxidoredutase NADP-N- quinona oxidoredutase Glutationa não enzimática a-Tocoferol (vitamina E) β- Caroteno Uratos Bilirrubina Flavonóides Albumina Ceruloplasmina Transferrina Tabela 1. Tabela resumo de oxidantes e antioxidantes. O conteúdo das enzimas AO em diferentes tecidos do corpo varia significativamente. Assim, seu nível e atividade nas células do tecido conjuntivo são várias vezes menores do que nos órgãos parenquimatosos. O nível de AO enzimático nas células está sob controle genético. Assim, um aumento no conteúdo do radical ânion superóxido ou peróxido de hidrogênio no citosol das células é acompanhado pela ativação da transcrição de genes que desencadeiam a síntese de cerca de 40 proteínas, incluindo catalase, superóxido dismutase, hidroperoxidase e endonuclease de reparo de DNA. Quase todas as influências que levam ao aumento da formação de ERO nas células induzem a síntese de AO enzimático. Sob condições de hipóxia e hiperóxia, isto é, condições que aumentam a formação de ERO, o nível de AOs enzimáticos no interior das células aumenta. Em condições normais, o conteúdo de AO enzimático é relativamente constante e depende pouco do sexo, havendo uma ligeira diminuição do seu nível com a idade; Ao mesmo tempo, o funcionamento normal do sistema AO do corpo e o conteúdo do AO enzimático são drasticamente perturbados em condições críticas (feridas, lesões, bem como queimaduras extensas na pele. Lesões graves são acompanhadas por uma violação do equilíbrio dinâmico de pro -sistemas oxidantes e antioxidantes, enquanto o conteúdo e a atividade dos antioxidantes endógenos, incluindo as enzimas AO, diminuem acentuadamente dentro e fora das células. Como já observado, a enzima chave da defesa AO é a superóxido dismutase (SOD), descoberta por McCord J. e Fridovich I. em 1969. A SOD pertence à classe das enzimas redox e está presente em quantidades significativas em quase todos os pró e eucarióticos. células. No corpo humano, a SOD é encontrada em todos os órgãos e tecidos, com concentrações intracelulares particularmente elevadas. No sangue, a enzima é encontrada em concentrações vestigiais quando administrada exogenamente e é excretada inalterada pelos rins em 5 a 10 minutos. Altas concentrações de SOD, comparáveis aos níveis intracelulares, são observadas no líquido broncoalveolar. No espaço extracelular, a enzima está presente em quantidades muito pequenas. Como consequência, no caso da formação de um grande número de radicais ânions superóxido no espaço extracelular, por exemplo, no foco da inflamação, o organismo fica praticamente indefeso contra os efeitos nocivos dos radicais. A principal função do SOD é acelerar a reação de dismutação dos radicais superóxido formados durante a oxidação biológica. A taxa de reação espontânea em valores de pH neutro não excede 7 x 105 Mˉ1 sˉ1 na presença de SOD, aumenta para 2 x 109 Mˉ1 sˉ1 e superior, ou seja, a enzima acelera a reação em três a quatro ordens de magnitude; . SOD possui várias isoformas que diferem na estrutura do centro ativo. A isoenzima contendo ferro é característica apenas de microrganismos; A enzima contendo manganês (Mn-SOD) está localizada nas mitocôndrias e é resistente ao cianeto. No volume total da atividade da superóxido dismutase, a participação do Mn-SOD é pequena e chega a cerca de 15%. Até 40% da SOD concentrada no núcleo das células vem da forma de manganês da enzima, que dismuta até 20% dos radicais superóxido produzidos no núcleo da célula. A forma cobre-zinco da enzima (Cu, Zn-SOD), sensível ao cianeto, é encontrada no citosol e no espaço intermembranar das mitocôndrias. Acredita-se que o átomo de cobre proporciona atividade catalítica e o átomo de zinco estabiliza a estrutura da isoforma citosólica da enzima. Tióis de baixo peso molecular, ao reduzirem o íon Cu2+ presente no centro ativo da SOD, ativam a enzima e aceleram a dismutação do oxigênio. Na maioria dos casos, a enzima consiste em duas subunidades idênticas (o peso molecular varia de 30-40 kDa), cada uma das quais contém um íon cobre cataliticamente ativo e um íon zinco, ligados por um ligante comum - histidina imidazol 61. O papel mais importante da SOD para o metabolismo celular é predeterminado não apenas pela ampla distribuição desta enzima, mas também por uma série de suas propriedades únicas: alta estabilidade térmica, resistência à ação de proteases, agentes desnaturantes e amplo pH ótimo. de atividade catalítica. A enzima é bastante estável em soluções - pode suportar aquecimento de até 100° C por um minuto e não perde atividade na faixa de pH de 2 a 12. A catalase é uma enzima contendo heme com peso molecular de 250 kDa. Assim como a SOD, a catalase está presente em todas as células do corpo, mas seu nível varia em diferentes tecidos. Altos níveis de catalase são encontrados nos glóbulos vermelhos, rins e fígado. Nestes últimos peroxissomos, a enzima representa até 40% da proteína total. A reação catalisada pela catalase é geralmente a seguinte. A velocidade da catálise é extremamente alta: uma molécula de catalase por segundo decompõe até 44.000 moléculas de peróxido de hidrogênio. A atividade da catalase depende da relação entre o número de ligações dissulfeto e o número de grupos sulfidrila na molécula da enzima, que participam da formação de seu centro ativo. Por causa disso, a catalase é sensível à ação dos venenos tiol. Devido ao grande peso molecular da molécula, a enzima praticamente não penetra nas membranas celulares. Este é um obstáculo ao uso de medicamentos catalase na prática. As enzimas AO SOD e catalase, funcionando juntas, inativam prontamente ROS, radical ânion superóxido e peróxido de hidrogênio formados durante o metabolismo celular normal, bem como durante intensificação significativa dos processos de peroxidação lipídica. No entanto, estas enzimas têm fraca actividade em relação aos peróxidos lipídicos formados durante as reacções em cadeia de peroxidação lipídica. A destruição desses produtos é realizada com a participação do sistema enzimático da glutationa. Sistema enzimático de glutationa. A glutationa peroxidase contendo selênio pode decompor eficazmente os hidroperóxidos lipídicos e o peróxido de hidrogênio. A sua afinidade pelo peróxido de hidrogénio é superior à da catalase, pelo que a primeira funciona de forma mais eficaz em baixas concentrações de peróxido, enquanto, ao mesmo tempo, a catalase desempenha um papel fundamental na proteção das células do stress oxidativo causado por altas concentrações de peróxido de hidrogénio. A glutationa peroxidase catalisa a reação de oxidação da glutationa com a formação de sua forma conjugada durante a reação, o peróxido de hidrogênio se decompõe em água; 2G-SH + H2O2 → G-S-S-G + 2H2O A enzima também catalisa a reação da glutationa reduzida com hidroperóxidos lipídicos, sendo estes últimos convertidos em hidroxiácidos graxos: 2G-SH + ROOH → G-S-S-G + R-OH+ H2O Junto com isso, a glutationa peroxidase é capaz de reduzir peróxidos de origem proteica e de ácido nucléico. A glutationa peroxidase está localizada no citosol (cerca de 70%) e nas mitocôndrias (20-30%) de todas as células de mamíferos. O sítio ativo da enzima contém quatro átomos de selênio ligados covalentemente para formar selenocisteína. A falta de selênio na dieta é acompanhada por uma diminuição da atividade da glutationa peroxidase nas células e, consequentemente, por uma diminuição da resistência do organismo aos danos oxidativos. Em casos graves, isto pode levar ao desenvolvimento de uma patologia de radicais livres semelhante à deficiência de vitamina E, que se caracteriza por obesidade, alterações necrobióticas no fígado e hemólise dos glóbulos vermelhos (doença de Keshan). Outra enzima que utiliza a glutationa para realizar a proteção AO é a glutationa transferase. A glutationa transferase é encontrada predominantemente no citosol das células do fígado humano e representa 2-4% da quantidade total de proteína citosólica. A enzima, ou mais precisamente, um grupo de enzimas, realiza a conjugação da glutationa reduzida com compostos hidrofóbicos e a redução de peróxidos orgânicos. A eliminação de derivados de xenobióticos lipofílicos, bem como de produtos LPO, incluindo hidroperóxidos de ácidos graxos, é realizada através de sua redução, substituição nucleofílica ou adição à molécula de glutationa: ROOH + 2G - SH → ROH +G - S - S - G + H 2O+G - SH →R- S - G + XH+ G - SH → HR - SG Ao contrário da glutationa peroxidase, para os quais os melhores substratos são hidroperóxidos hidrofílicos com tamanho de molécula pequena, a glutationa transferase não interage com o peróxido de hidrogênio, mas reduz hidroperóxidos hidrofóbicos com grande volume de molécula: hidroperóxidos de ácidos graxos poliinsaturados - ácidos linolênico e araquidônico, também como fosfolipídios. Além disso, a enzima restaura hidroperóxidos de mononucleotídeos e DNA, participando assim de seu reparo. A glutationa transferase é um componente importante da defesa da AO, garantindo a remoção de muitos metabólitos do estresse oxidativo. A glutationa reduzida é necessária para o funcionamento normal das enzimas dependentes da glutationa. A manutenção do seu nível suficiente é realizada através da síntese, devido à sua dessorção das ligações com proteínas, ou através da recuperação da oxidação da forma. Esta reação é realizada pela enzima glutationa redutase, uma flavoproteína. G-S-S-G + NADPH + H+ → 2G - SH + NADP + O sistema glutationa redutase permite reabastecer rapidamente o pool de glutationa reduzida nos tecidos. A principal fonte de NADPH para esta reação é o ciclo das pentoses de oxidação da glicose, enquanto as desidrogenases dependentes de NADPH do ciclo das pentoses são ativadas pela glutationa oxidada. A diminuição do conteúdo de glutationa reduzida nos tecidos cria condições favoráveis para o bloqueio de grupos SH que fazem parte de enzimas e proteínas estruturais da célula. Acredita-se que a glutationa seja o principal agente que protege as enzimas tiol da oxidação. As enzimas tiol incluem muitas oxidoredutases, transferases, hidrolases, liases e ligases. A oxidação dos grupos SH perturba o funcionamento da glicose-6-fosfato desidrogenase, lactato desidrogenase, succinato desidrogenase, ATPase, monoamina oxidase, xantina oxidase, glutationa redutase e catalase. Os fatos acima confirmam a opinião de que os compostos tiol desempenham o papel mais direto e amplo no mecanismo de funcionamento da ligação enzimática do sistema AO, desempenhando não apenas um efeito anti-radical, mas também um efeito antiperóxido. Compostos tiol solúveis em água de baixo peso molecular: glutationa e ergotioneína. O primeiro deles é um componente do sistema redox tiol dissulfeto não proteico e é um tripéptido formado pelos aminoácidos cisteína, ácido glutâmico e glicina. A glutationa existe em duas formas - reduzida (até 97% do total) e oxidada. Juntamente com outros compostos contendo enxofre, a glutationa é um inibidor de ERO e estabiliza as membranas celulares. Desempenha um papel importante na neutralização do radical hidroxila formado na reação de Fenton ou como resultado da ridiólise da água sob a influência da radiação ionizante. 1.6 Mecanismos patogenéticos de distúrbios que se desenvolvem quando o equilíbrio dos sistemas antioxidante e pró-oxidante é perturbado Um desequilíbrio entre os sistemas antioxidante (AOS) e pró-oxidante (POS) causa o desenvolvimento do estresse oxidativo (OS). O efeito tóxico das ERO se manifesta nas condições do sistema operacional, que é acompanhado por uma forte intensificação dos processos de radicais livres nos tecidos. Este é o elo patogenético mais importante no desenvolvimento de muitos processos inflamatórios, lesões por radiação, doenças cardiovasculares, câncer, intoxicações químicas e outras. O mecanismo de geração de ERO em muitas condições patológicas é geral. Algumas características distintivas podem ser identificadas apenas nos estágios iniciais. Assim, durante os processos inflamatórios, o fator desencadeante para a intensificação dos processos de radicais livres é uma explosão respiratória durante a hipóxia, uma perturbação principalmente do sistema respiratório tecidual e durante lesões químicas, ativação do sistema de oxidação microssomal; Assim, as razões que causam a intensificação dos processos de radicais livres podem ser diferentes, mas as mudanças no nível molecular são do mesmo tipo e os processos de geração de ERO estão inter-relacionados. Alguns antioxidantes sob condições OS podem atuar como pró-oxidantes. Nas condições de OS, o potencial de recuperação da célula aumenta devido a substratos e coenzimas em estado reduzido, o que leva à diminuição do pH nas áreas de isquemia do tecido cardíaco e cerebral. Isto cria condições para aumentar o conjunto de “formas ativas” de metais de valência variável. Sob condições de maior geração de EROs, eles podem participar de reações associadas à geração de produtos radicais. Assim, na presença de Fe/Cu e O2, os tióis (RSH) são fontes de radicais, RS , O2 -, H2O2 e OH , radicais NADPH NAD(P) , o ácido ascórbico é um radical semidesidrascorbato. Um aumento no nível de ERO está associado a uma intensificação dos processos de destruição oxidativa de lipídios, proteínas, ácidos nucléicos e carboidratos. É a intensificação desses processos a principal causa do dano citotóxico aos tecidos. Como já indicado, a razão para o acúmulo de espécies reativas de oxigênio são distúrbios no metabolismo oxidativo normal das células durante a patologia (desvio pelo oxigênio das vias de transporte elétrico nas mitocôndrias e no sistema de oxidação macrossomal devido ao “vazamento” de elétrons, transformação de vias de desidrogenase em vias de oxidase, reações de autooxidação, por exemplo, catecolaminas, etc.). Em concentrações superiores às fisiológicas, todos estes compostos são altamente tóxicos para os sistemas biológicos em todos os níveis, desde o molecular-celular até ao organismo. A capacidade de causar danos oxidativos e destruição de componentes de sistemas vivos, causada por derivados ativos de oxigênio, é designada como “toxicidade de oxigênio”. Os radicais livres de oxigênio são compostos químicos instáveis e reagem facilmente com biomoléculas, causando sua modificação ou destruição. Os objetos de reações oxidativas não específicas e não autorizadas envolvendo oxigênio e seus derivados quimicamente ativos podem ser moléculas de diversas naturezas químicas. As ROS são capazes de quebrar qualquer ligação de hidrocarbonetos e destruir facilmente compostos de alto peso molecular: ácido hialurônico, proteoglicanos, colágeno, imunoglobulinas. Na presença de íons ferrosos, as ROS convertem a oxiemoglobina em metemoglobina. A oxidação de grupos funcionais de substâncias biologicamente ativas causa degradação de proteínas estruturais e lipídios de membranas celulares e ácidos nucléicos, inibição de enzimas, alterações na estrutura e propriedades de hormônios e seus receptores. O resultado da interação de derivados ativos de oxigênio com a molécula de DNA é a modificação estrutural das bases nitrogenadas, a decomposição do anel desoxirribose de cinco membros, bem como a clivagem da estrutura açúcar-fosfato, o que acaba levando à fragmentação de este polímero. Foi provado que a produção excessiva de ERO, em particular, radical ânion superóxido, oxigênio singleto e radical hidroxila, pode causar uma série de aberrações cromossômicas e mutações de vários genes em linfócitos humanos. A interação das ERO com proteínas leva a alterações estruturais neste tipo de biomolécula, consistindo na modificação de grupos laterais de resíduos de aminoácidos, fragmentação de cadeias polipeptídicas e formação de ligações covalentes dentro e entre moléculas. A tolerância das proteínas às ROS depende da sua composição de aminoácidos cíclicos e os aminoácidos contendo enxofre são mais sensíveis aos danos dos radicais livres; Dos 20 aminoácidos essenciais, os mais vulneráveis às ERO, em particular ao oxigénio singleto, são a histidina, o triptofano, a metionina, a tirosina e a cisteína. Uma característica distintiva do dano oxidativo aos lípidos nas membranas celulares é que o oxigénio molecular pode acumular-se na região hidrofóbica dos seus resíduos de ácidos gordos. Como resultado disso, bem como do arranjo próximo e paralelo dos resíduos de ácidos graxos vizinhos em relação uns aos outros, são criadas condições para o desenvolvimento de danos oxidativos como reações em cadeia. Devido ao papel fundamental dos hidroperóxidos orgânicos nesta expansão, este processo é denominado peroxidação lipídica. 2. Radicais livres (peroxidação lipídica) Os principais substratos do LPO são os ácidos graxos superiores poliinsaturados (HFA), localizados na estrutura dos fosfolipídios da membrana. Em diferentes estágios de peroxidação de AGV, conjugados de dieno e trieno, formam-se peróxidos de BFA (R-OO) ˙), Hidroperóxidos de AGV (R-OO), endoperóxidos, malondialdeído e novos radicais livres. O catalisador mais forte para o processo são os íons metálicos (Fe2+). O processo pode ser interrompido pela formação de produtos que não contenham radicais livres. Os processos LPO podem ser condicionalmente divididos em três estágios sucessivos, ou fases de desenvolvimento: processos de iniciação da cadeia, processos de desenvolvimento de reações em cadeia e terminação da cadeia. Na fase de nucleação da cadeia, sob a influência dos radicais livres do oxigênio, da radiação ionizante, da irradiação ultravioleta e de uma série de substâncias químicas relacionadas aos pró-oxidantes, ocorre a formação de radicais orgânicos (R). O2 + RH→R ˙ + HO2 O2 + RH→R ˙ +HO2 ELE ˙ + RH → R ˙ + H2O Na próxima etapa, o radical interage rapidamente com o oxigênio, que atua como aceptor de elétrons. Como resultado, ocorre a formação do radical peróxi (RO2), que ataca os lipídios insaturados. O surgimento de peróxidos orgânicos e de um novo radical (R) como resultado desta reação contribui para a continuação de reações oxidativas que adquirem caráter em cadeia: R + O2 → RO2;+ RH → ROOH + R Os peróxidos orgânicos (ROOH) são incluídos no processo de geração de radicais na presença de metais de valência variável (cobre, cobalto, manganês, ferro), forma-se um radical alcóxi de reação: ROOH + Me n+→RO + OH ˉ + Me (n+1)- Alguns dos radicais orgânicos resultantes interagem entre si, resultando na formação de moléculas inativas, o que interrompe o curso das reações de oxidação dos radicais livres: RO2 + R → ROOR Os hidroperóxidos lipídicos são capazes de sofrer transformações oxidativas não radicais, o que leva à formação de produtos de peroxidação lipídica primária (conjugados de dieno, dialdeídos), intermediários (bases de Schiff) e finais, além de álcoois, cetonas e aldeídos. A quebra de reações em cadeia de peroxidação é possível através da interação de radicais com sistemas enzimáticos especializados, bem como com uma série de substâncias de baixo peso molecular que formam coletivamente o componente bioquímico do sistema antioxidante do corpo. A ativação excessiva do LPO pode ocorrer como resultado de mudanças repentinas no regime de oxigênio da célula. Nesse caso, a hiperóxia causa um aumento temporário nos processos de peroxidação lipídica, e a hipóxia persistente leva a um acúmulo semelhante a uma avalanche de produtos tóxicos de peroxidação. A intensidade excessiva das reações de peroxidação lipídica se desenvolve sob a influência de influências externas, que incluem radiação ionizante e ultravioleta, bem como uma série de substâncias químicas relacionadas aos pró-oxidantes. A produção de ERO e, consequentemente, a intensidade dos processos de peroxidação lipídica nas células aumentam acentuadamente como uma reação inespecífica quando o corpo é exposto a diversos fatores de estresse de natureza química, física e biológica. Em última análise, todos estes efeitos podem levar à tensão e subsequente descompensação dos mecanismos de defesa antioxidante do corpo e ao desenvolvimento de stress oxidativo, manifestado a nível celular, tecidual e do organismo. Processos patológicos típicos, como hipóxia e inflamação, característicos e em desenvolvimento na maioria das doenças somáticas e infecciosas, lesões e feridas graves, são sempre acompanhados pela formação excessiva de ROS e produtos de peroxidação lipídica A ativação prolongada dos processos de peroxidação lipídica é acompanhada pelo desenvolvimento da síndrome de peroxidação lipídica. Com a ativação excessiva da peroxidação lipídica, quando uma parte significativa dos fosfolipídios da membrana sofre degradação oxidativa, a fase lipídica das membranas torna-se mais rígida. Isto limita a mobilidade conformacional da cadeia polipeptídica, como resultado da redução da atividade funcional de enzimas, receptores e proteínas formadoras de canais incorporadas nas membranas, o que por sua vez impede a remoção de Ca+2 do sarcoplasma e garante a efeito prejudicial do cálcio nas organelas celulares. Até o momento, foi comprovada a capacidade dos metabólitos da LPO de inibir a K+/Na+-ATPase da membrana plasmática e causar citólise de hepatócitos com desenvolvimento de fermentemia. Os produtos LPO perturbam o funcionamento normal dos canais de cálcio do tipo L, que é acompanhado por distúrbios eletrolíticos nas células do miocárdio. Seu excesso leva a uma mudança no metabolismo do ácido araquidônico, como resultado da redução de um dos compostos resultantes (ácido 15-hidroaraquidônico). β- estimulação adrenérgica devido à inibição deste tipo de receptores adrenérgicos. Os produtos dessas reações danificam as membranas dos eritrócitos, oxidam a albumina sérica e interrompem a síntese de ácidos nucléicos nas células. Espécies de oxigênio ativadas e produtos de peroxidação lipídica danificam o ácido hialurônico, os proteoglicanos, o colágeno e as imunoglobulinas. A geração de espécies de oxigênio ativadas que não é compensada pelo sistema antioxidante pode causar um efeito prejudicial no endotélio e na membrana basal vascular, direta ou indiretamente, através da inativação dos sistemas antioxidantes. Grânulos de lipofuscina morfologicamente detectáveis, constituídos por depósitos de lipoperóxidos e proteínas insolúveis, são produtos da peroxidação lipídica. ROS e produtos de peroxidação lipídica contribuem para a produção e liberação excessiva de uma série de citocinas pró-inflamatórias (fator de necrose tumoral, interleucina-1, interleucina-6) e mediadores inflamatórios (histamina, bradicinina, serotonina), derivados do ácido araquidônico - leucotrienos, prostaglandinas e tromboxanos. A produção excessiva de EROs e produtos de peroxidação lipídica causa uma ativação acentuada do sistema complemento ao longo da via alternativa com liberação de anafilatoxina. 2.1 Participação de produtos ROS e LPO na patogênese de doenças humanas A maioria ou quase todas as condições patológicas, cujo curso é acompanhado pela participação de ERO, são caracterizadas pelo chamado estado de estresse oxidativo, caracterizado pela intensificação da produção dessas substâncias. Ressalta-se que o estado de estresse oxidativo é característico não apenas de doenças, mas também ocorre em estados fisiológicos próximos ao extremo - intenso estresse físico e psicoemocional, excesso de trabalho, bem como durante o envelhecimento do corpo. Existe uma “teoria radical livre do envelhecimento”, seu criador D. Harman, que delineou pela primeira vez as principais disposições da teoria em 1955. A teoria de Harman é baseada em um sistema de argumentos associados à capacidade dos radicais livres de danificar macromoléculas de forma inespecífica, como como DNA, lipídios e proteínas, e o processo de envelhecimento está associado ao acúmulo desses danos no organismo. No quadro desta teoria, explica-se não só o envelhecimento do corpo, mas também uma vasta gama de processos patológicos associados ao envelhecimento: doenças cardiovasculares, disfunções cerebrais relacionadas com a idade, imunossupressão, carcinogénese e outros tipos de patologia. Em todas as doenças em que uma reação inflamatória está presente como componente obrigatório, o papel principal nos danos às células e tecidos do corpo é desempenhado pelas ERO produzidas pelos leucócitos, principalmente nertrófilos. Embora as formas mais destrutivas de ERO geradas pela mieloperoxidase atuem nos fagolisossomos do leucócito, no local da inflamação os neutrófilos são capazes de secretar essa enzima no espaço extracelular. A destruição de um agente infeccioso durante o desenvolvimento do processo inflamatório também pode ser alcançada à custa de danos colaterais aos próprios tecidos no local da inflamação, enquanto os danos aos tecidos dependentes de oxigênio são diretos e indiretos. Este último é realizado quando as ROS influenciam moléculas reguladoras e efetoras, por exemplo, proteinases e inibidores de proteinases. As ERO podem ativar diretamente formas latentes de colagenase secretadas pelos neutrófilos, bem como inativar o inibidor universal da proteinase - α 2-macroglobulina e inibidor específico de serina proteinases - α ı- antitripsina. As ERO, geradas intracelularmente e também penetrando através da membrana celular, são gatilhos para a indução da apoptose. O mecanismo dos radicais livres de danos às membranas plasmáticas, mitocondriais e nucleares, ao genoma nuclear e mitocondrial, às lipoproteínas sanguíneas leva a danos aos vasos sanguíneos e às barreiras histohemáticas, que desempenham um papel importante, muitas vezes decisivo, na patogênese das doenças inflamatórias mais comuns. , natureza tóxica e autoimune. O papel patogenético das ERO foi agora identificado para aproximadamente centenas de doenças humanas. Isso ocorre na patologia cardiovascular - doença coronariana, infarto do miocárdio, obstrução arterial aguda de um segmento do membro, danos isquêmicos e de reperfusão nos rins, cérebro e outros tecidos, no desenvolvimento de catarata e aterosclerose. Foi demonstrado que a formação de placas ateroscleróticas na íntima vascular ocorre com a participação ativa de radicais livres interagindo com ácidos graxos poliinsaturados. A patogênese do lúpus eritematoso sistêmico baseia-se no aumento da sensibilidade dos ácidos nucléicos (em particular, do DNA nuclear) aos efeitos prejudiciais dos radicais livres. Um fator predisponente para a doença é uma dieta com excesso de ácidos graxos poliinsaturados. As ERO desempenham um papel importante na patologia broncopulmonar, tanto em doenças agudas como crónicas (por exemplo, enfisema, asma e bronquite crónica). Os radicais livres são extremamente importantes na patogênese da síndrome do desconforto respiratório. São as ERO produzidas pelos neutrófilos do sangue que desempenham o papel principal nos danos à barreira endotelial-alveolar, no desenvolvimento de edema intersticial e no quadro clínico completo desta síndrome. A estimulação excessiva e prolongada dos sistemas enzimáticos responsáveis pela geração de radicais ativos dos fagócitos está subjacente aos mecanismos de formação da doença ocupacional da poeira e suas complicações, como a colagenose sistêmica e a degeneração maligna dos elementos celulares dos pulmões e da pleura. Um estudo retrospectivo de longo prazo realizado em 23 países sob os auspícios da OMS revelou uma dependência direta da incidência de certos tipos de neoplasias malignas de pulmões, mama, ovários e reto da quantidade de gordura animal consumida e da intensidade da peroxidação lipídica processos no corpo. Numerosos estudos indicam que os processos de oxidação de radicais livres estão subjacentes à patogénese de muitas doenças hepáticas crónicas, e a formação excessiva de ERO e produtos de peroxidação lipídica manifesta-se nas fases iniciais do processo. O efeito hepatotóxico dos radicais de oxigênio se manifesta nas membranas celulares, que são o principal local de síntese do radical ânion superóxido. O aumento da produção de ERO como resultado da indução da xantina oxidase pelo interferon ocorre mesmo durante a infecção viral. Os radicais de oxigênio danificam o aparelho de membrana do hepatócito, destroem a camada lipídica de suas membranas e também danificam proteínas - enzimas do sistema monooxigenase. A gravidade e o prognóstico da doença hepática crônica estão intimamente relacionados ao estado funcional do sistema antioxidante dos hepatócitos. A ativação excessiva de LPO é a causa do aparecimento e progressão da hipertensão. Foi estabelecido que sob a influência de EROs e produtos de peroxidação lipídica, a permeabilidade das membranas celulares e a atividade da Na+/K+-ATPase são perturbadas, o que leva ao acúmulo de cálcio ionizado nas células. Além disso, os produtos secundários de LPO podem afetar diretamente o tônus dos vasos arteriais. Além do efeito vasoconstritor, os radicais de oxigênio ativam o trabalho dos fibroblastos do tecido conjuntivo, resultando no desenvolvimento de fibrose arteriocapilar, ou seja, alterações irreversíveis nos vasos sanguíneos. O estresse oxidativo quase sempre se desenvolve em doenças infecciosas. A formação excessiva de radicais livres de oxigênio é um dos principais mecanismos na patogênese da gripe. A geração de ERO determina a mutagênese e a atividade proteolítica do vírus influenza, o efeito citopático da infecção viral, a destruição da rede capilar e das paredes capilares, o desenvolvimento de pneumonia confluente e edema pulmonar. Danos tóxicos aos neurônios por ERO, produtos de peroxidação lipídica e estresse oxidativo são considerados o principal mecanismo em doenças neurodegenerativas. Uma das razões para o desenvolvimento da esclerose múltipla podem ser distúrbios mitocondriais acompanhados de produção excessiva de ERO. O oxigênio ativo na esclerose múltipla tem um efeito prejudicial direto no sistema nervoso central e é um fator causal no desenvolvimento da doença. O dano oxidativo leva à morte das células nervosas e gliais, que se manifesta pela desmielinização e pela formação de focos de gliose no sistema nervoso central. Em 37% das pessoas com doença de Parkinson, existe um defeito no primeiro complexo da cadeia respiratória nas células da substância negra. A acumulação gradual de metabolitos de oxigénio, produtos de peroxidação lipídica, metaloproteínas e o desenvolvimento de stress oxidativo em várias partes do sistema nervoso central são de particular importância na doença de Alzheimer e na coreia de Huntington. A acumulação de lipofuscina nas células do SNC sob estas condições correlaciona-se com o nível de enzimas antioxidantes no tecido cerebral. As ROS e os produtos da peroxidação lipídica têm um efeito destrutivo direto nos órgãos internos e levam ao desenvolvimento de falência múltipla de órgãos após lesões graves e queimaduras extensas. A radiólise da água, a formação de radicais livres de oxigênio e vários metabólitos da peroxidação lipídica estão na base da patogênese das lesões agudas e crônicas por radiação em humanos. 2.2 Diagnóstico de processos de peroxidação Não há sinais clínicos claros de ativação excessiva da oxidação dos radicais livres no corpo humano. O desenvolvimento de estresse oxidativo em um paciente é acompanhado por: sinais de dificuldade e insuficiência respiratória circulação sanguínea (ocorre frequentemente um tipo hiperdinâmico de circulação sanguínea); agitação psicomotora; um aumento de temperatura superior a 37,5° C; reação de dor pronunciada. Os autores consideram que os sinais laboratoriais de estresse oxidativo são: aumento do número de leucócitos no sangue (> 10 mil) sem mudança de banda em futuro próximo após lesão (10-12 horas); níveis aumentados de alanina e transaminases aspárticas no sangue; aumento da concentração plasmática de glicose > 7,0 mmol/l; acidose metabólica descompensada (pH< 7,2); Para diagnosticar laboratorialmente a gravidade do estresse oxidativo e determinar as indicações para o uso de medicamentos com propriedades antioxidantes, é aconselhável avaliar o estado do sistema antioxidante, bem como a intensidade das reações de radicais livres (processos LPO) no corpo do paciente . A avaliação do estado do sistema antioxidante geralmente inclui a determinação da atividade dos principais antioxidantes enzimáticos (atividade da SOD, catalase e glutationa peroxidase dos eritrócitos), bem como o conteúdo de ácido ascórbico, tocoferol e selênio no sangue. A avaliação instrumental da gravidade das reações de radicais livres envolve a determinação de produtos de reação de peroxidação em amostras de sangue ou tecidos, que incluem conjugados de dieno, malondialdeído, bases de Schiff e espécies reativas de oxigênio. A combinação destes indicadores permite avaliar tanto o estado dos principais elos do sistema AO do corpo, ou seja, a sua capacidade de prevenir reações excessivas de natureza radicalar livre, como a real intensidade das reações de LPO e síntese de ROS. Com base nisso, esses exames podem ser considerados critérios laboratoriais para diagnóstico de estresse oxidativo. O diagnóstico laboratorial da intensidade da peroxidação lipídica é necessário para avaliar a atividade AO de um determinado medicamento, pois com base nos sinais clínicos é impossível avaliar adequadamente o efeito do medicamento AO. Observe que usar apenas um dos testes acima não fornece informações completas. A dinâmica dos indicadores da intensidade das reações dos radicais livres e do conteúdo de AO no corpo às vezes tem direção oposta. Em alguns casos, as flutuações no conteúdo dos produtos da reação dos radicais livres não são acompanhadas por alterações nos níveis de espécies reativas de oxigênio e vice-versa. Uma forte intensificação das reações dos radicais livres e o acúmulo dos produtos dessas reações no sangue podem não levar a uma alteração no conteúdo e na atividade dos principais componentes do sistema AO. Para uma interpretação clínica fiável e adequada dos resultados obtidos, não só o valor absoluto dos indicadores é de fundamental importância, mas a sua dinâmica no contexto das medidas de tratamento. Critérios laboratoriais para diagnóstico de estresse oxidativo Avaliação do estado do sistema antioxidante do paciente Determinação da intensidade das reações dos radicais livres Conteúdo e atividade de enzimas antioxidantes em eritrócitos ou outras células disponíveis para estudo: Superoxido dismutação Catalase Glutationa peroxidaseConteúdo no plasma sanguíneo (soro) de produtos primários, secundários e intermediários da peroxidação lipídica: Conjugado de dieno Malondialdeído Bases de SchiffConteúdo de antioxidantes de baixo peso molecular no sangue: Ácido ascórbico Tocoferol SelênioConteúdo de formas reativas (radicais) de oxigênio no sangue: Radical ânion superóxido Oxigênio singleto Nível total de radicais Cada um dos indicadores propostos tem suas vantagens e desvantagens. Os principais requisitos para métodos laboratoriais são precisão, facilidade de implementação e boa reprodutibilidade. A facilidade de preparação das amostras para teste e a possibilidade de armazenamento são desejáveis, o que torna possível não realizar todos os testes ex tempore. O método mais utilizado na pesquisa experimental e na prática clínica para determinação do malondialdeído na reação com o ácido tiobarbitúrico não é um indicador preciso que reflita o curso dos processos de peroxidação lipídica, uma vez que durante a reação o ácido tiobarbitúrico interage com substâncias não lipídicas contendo um grupo ceto, por exemplo, com glicose. A interação do ácido tiobarbitúrico com o malondialdeído, inicialmente contido nos sistemas lipídicos, desempenha um papel insignificante em termos quantitativos. Porém, há quase trinta anos, o método tem sido o mais utilizado pela maioria dos pesquisadores no estudo de processos de peroxidação lipídica em diversos sistemas biológicos. Suas vantagens incluem simplicidade e relativa velocidade de execução, além de boa reprodutibilidade dos resultados. Outros métodos comuns para avaliar a intensidade das reações de LPO são determinar o nível de produtos primários e intermediários dessas reações, isto é, conjugados de dieno e bases de Schiff. Os métodos são precisos e relativamente fáceis de reproduzir. Suas desvantagens incluem a impossibilidade de armazenamento das amostras selecionadas, portanto os estudos devem ser realizados imediatamente ou meia hora após sua coleta. Um indicador mais preciso e adequado da intensidade e gravidade das reações dos radicais livres e, consequentemente, do estado do sistema antioxidante do corpo é a determinação de ROS - os principais precursores das reações de LPO; a medição rápida e precisa do nível no sangue total é realizada pelo método luminescente. Dependendo do tipo de fósforo utilizado (luminol, lucigenina, etc.), é possível determinar tanto os tipos individuais de ROS quanto sua quantidade total. O método é rápido e pode ser recomendado para diagnósticos expressos. A técnica permite armazenar amostras de sangue heparinizadas para pesquisa por até 12 horas a uma temperatura de +4˚C, ou seja, em uma geladeira doméstica comum. Principais indicadores que caracterizam o sistema oxidante-antioxidante. A intensidade do LPO é avaliada pela concentração no sangue e outros fluidos biológicos de produtos de reação intermediários e finais. O malondialdeído (MDA) é o produto final do LPO. A concentração normal no sangue é de 2,5-6,0 µM/l. Depende do método de determinação, portanto cada laboratório possui seu próprio intervalo de referência. Um aumento na concentração é evidência de aumento de LPO e interrupção da proteção antioxidante. Determinação de malondialdeído no sangue pelo método fluorimétrico (Fedorova T. N., Koryatsiva T. S., Larsky E. G.) O princípio do método: o ácido tiobarbitúrico (TBA) em ambiente ácido reage com dialdeídos de baixo peso molecular (principalmente ácido malônico) para formar um complexo de cor rosa. Valores padrão: 3,7±0,12 unidades. Modificação da determinação de produtos de peroxidação lipídica na reação com ácido tiobarbitúrico (Korobeinikov E. N.) O princípio do método: quando aquecido em ambiente ácido, parte dos produtos LPO pertencentes à classe dos hidroperóxidos se decompõe com a formação do malondialdeído, cuja interação de uma molécula com duas moléculas de ácido tiobarbitúrico leva à formação de um complexo colorido. Valores padrão: 3,69±0,14nmol/l. Determinação de peróxidos lipídicos na reação com ácido tiobarbitúrico (Gavrilov V. B., Gavrilova A. R., Mazhul L. M.) Princípio do método: o ácido tiobarbitúrico reage com o malondialdeído, que se forma durante a peroxidação de ácidos graxos insaturados com 2-3 ligações dieno, para formar um produto rosa com absorção máxima em 535 nm. Valores padrão: 100-120 nmol/l. Determinação de conjugados de dieno no plasma sanguíneo por absorção UV extratos de heptano e isopropanol (Gavrilov V.B., Gavrilova A. R., Khmara N. F.) Princípio do método: o método baseia-se na medição da intensidade de absorção na região de 223-234 nm, causada por estruturas dieno conjugadas (previamente extraídas do plasma) que surgem durante a formação de hidroperóxidos de ácidos graxos poliinsaturados. Determinação espectrofotométrica do conteúdo de hidroperóxidos de acila (conjugados de dieno) no plasma sanguíneo (soro). O princípio do método baseia-se na determinação do conteúdo de produtos primários da peroxidação lipídica no sangue pela absorção de um fluxo de luz monocromático por um extrato lipídico na região ultravioleta do espectro (233 nm). Vitaminas com propriedades antioxidantes. A vitamina A (retinol) é uma vitamina lipossolúvel encontrada em produtos de origem animal na forma de A e A2, em plantas na forma de pró-vitamina - caroteno, sendo o mais ativo o beta-caroteno. A concentração sérica normal é 1,05-2,27 µM/L ou 300-650 µg/L. A deficiência de vitamina A, que pode ocorrer não apenas com deficiência de retinol ou caroteno nos alimentos, mas também devido à absorção prejudicada devido a doenças intestinais, bem como com pequena quantidade de gordura e proteína nos alimentos, contribui para o aumento do LPO. O epitélio de vários órgãos e do olho sofre mais (recuperação lenta da púrpura visual - rodopsina). Isto se manifesta por aumento da suscetibilidade a doenças broncopulmonares, infecções do trato urinário, membranas mucosas dos olhos, pele e diminuição da acuidade visual no escuro (“cegueira noturna”). A vitamina E (tocoferóis) é um grupo de substâncias entre as quais o alfa-tocoferol tem maior atividade. Vitamina lipossolúvel que atua em conjunto com a vitamina A. No soro sanguíneo e nas células, a maior parte está associada às lipoproteínas. Determinação do nível de alfa-tocoferol no plasma sanguíneo. Princípio do método: O alfa-tocoferol plasmático é extraído com uma mistura de etanol-hexano. A fase hexânica selecionada é evaporada. Ao resíduo resultante é adicionado cloreto férrico, que é capaz de ser reduzido do estado trivalente para o estado divalente sob a influência de antioxidantes (e principalmente alfa-tocoferol). A quantidade de alfa-tocoferol é avaliada pelo teor de ferro reduzido, que é determinado pela reação com alfa-2-, alfa-2-dipiridil, acompanhada pela formação de um complexo vermelho. A concentração normal no soro sanguíneo é de 5-20 mg/l. O grau de fornecimento de vitamina E ao corpo pode ser caracterizado pela determinação direta de sua concentração no sangue e indiretamente pela determinação da resistência ao peróxido dos eritrócitos. A resistência aos peróxidos dos eritrócitos (PRE) é um indicador do fornecimento de antioxidantes às membranas, principalmente vitamina E, e de sua resistência aos efeitos prejudiciais dos peróxidos. O valor normal é de até 10%. O aumento indica proteção antioxidante insuficiente das membranas celulares, alta suscetibilidade à peroxidação lipídica e labilidade. Existem flutuações sazonais no ERP com aumento na porcentagem de hemólise de peróxido na primavera. . A vitamina C (ácido ascórbico) é uma vitamina solúvel em água que, ao contrário dos animais, não é sintetizada no corpo humano e vem apenas com os alimentos. Previne a oxidação das vitaminas A e E, restaura formas oxidadas de enzimas e diversos substratos e participa de um grande número de reações metabólicas. Regula a permeabilidade da parede vascular, aumenta significativamente a resistência do organismo a infecções e é necessária para a formação de colágeno, cicatrização de feridas e queimaduras. Concentração normal: no sangue -6-20 mg/l, na urina -20-30 mg/dia. Necessidade diária - 45-60 mg; aumenta significativamente com estresse físico e neuropsíquico, baixas e altas temperaturas, radiação, tabagismo (1 cigarro destrói 25 mg de vitamina C), gravidez e lactação, doenças crônicas do trato gastrointestinal, uso de antibióticos e sulfonamidas. O ácido ascórbico não é recomendado para pacientes com câncer durante radiação e quimioterapia. A falta de vitamina C nos alimentos contribui para o aumento da peroxidação lipídica, distúrbios metabólicos de muitas substâncias, funções dos sistemas nervoso e endócrino, diminuição da resistência a infecções e se manifesta por fraqueza, aumento da fadiga, irritabilidade, dores nos membros e outros sintomas inespecíficos. A deficiência de vitaminas a longo prazo causa o desenvolvimento do escorbuto. Enzimas antioxidantes. A superóxido dismutase (SOD) é uma enzima que catalisa a reação de neutralização do radical superóxido O2. É a principal enzima da defesa antirradicalar intracelular. A atividade normal nos eritrócitos (método de recuperação NBT) é 1,04 ± 0,05 arb. unidades/mg de hemoglobina. Os valores absolutos variam dependendo dos reagentes utilizados. O mais acessível para análise é o SOD eritrocitário contendo CuZn. Determinação da atividade de SOD eritrocitária pelo método de R. Fried. Princípio do método: o método baseia-se na determinação do grau de inibição da reação de redução do nitroazul tetrazólio por radicais superóxido sob a influência da SOD eritrocitária. O estudo da atividade da SOD é realizado à temperatura ambiente. O sangue deve ser coletado com heparina. A enzima pode permanecer estável por até 1 semana quando armazenada a +4 C. A atividade enzimática é expressa em unidades arbitrárias (unidades arbitrárias). 50% de inibição do processo de redução do nitroazul tetrazólio em comparação com a amostra de controle é considerada 1 arb. unidades Valores padrão: 1,04±0,3/mg de hemoglobina. Durante o período inicial de aumento da formação de radicais livres, a atividade da SOD aumenta para neutralizá-los. Se o processo de formação de radicais continuar a se intensificar, então, em determinado estágio, as reservas compensatórias se esgotam e a atividade da SOD diminui abaixo do normal, o que indica descompensação desse mecanismo protetor. Um aumento na atividade da SOD é observado no envenenamento por haxixe, tetracloreto de carbono, amitriptilina, compostos organofosforados, ácido acético, septicemia, tuberculose focal, leucemia, distrofia muscular de Duchenne, fibrose cística, talassemia, infarto do miocárdio e doença mental. Um aumento na atividade da SOD sem a correspondente ativação ou ausência de outras enzimas antioxidantes - catalase (hipo-, acatalasemia), peroxidases é uma alteração desfavorável ao organismo devido ao acúmulo de peróxido de hidrogênio - produto da reação da superóxido dismutase. Observa-se diminuição da atividade da SOD em caso de envenenamento por monóxido de carbono, compostos de chumbo, cádmio, com septicopemia, com cardiopatia isquêmica, aterosclerose cerebral, com epilepsia, em recém-nascidos com síndrome do desconforto respiratório, com retinopatia, em gestantes com toxicose tardia (teste diagnóstico para determinar a hipóxia intrauterina do feto). Baixos níveis de atividade enzimática devem ser considerados um sinal prognóstico desfavorável, indicando diminuição da resistência inespecífica do organismo. É aconselhável determinar a atividade do SOD com estudo simultâneo de outros componentes do AOD e indicadores de LPO. A catalase é uma enzima que decompõe o peróxido de hidrogênio em oxigênio e água. A quantidade máxima é encontrada nos glóbulos vermelhos. Possui função protetora antioxidante específica contra células endoteliais. A atividade normal é de 18,4 - 25,0 µU/eritrócito. Na fase compensatória inicial da formação do radical ocorre aumento da atividade enzimática e na fase de descompensação ocorre diminuição. Com a idade, a atividade enzimática diminui. Determinação da atividade da catalase pelo método de E. Beutler. Princípio do método: o método baseia-se na determinação da taxa de decomposição do peróxido de hidrogênio em mM/min espectrofotometricamente em um comprimento de onda de 230. O etanol é adicionado para estabilizar o hemolisado e decompor o complexo catalase-peróxido de hidrogênio. Determinação da atividade da catalase sanguínea pelo método de Bach e Zubkova. Princípio do método: o método baseia-se na determinação da quantidade de peróxido de hidrogênio que é destruído pela enzima em 30 minutos. No experimento, determina-se a quantidade de peróxido de hidrogênio não destruído durante a reação da catalase, determina-se a quantidade de peróxido retirada em uma amostra com a enzima inativada por fervura; O peróxido de hidrogênio é titulado com permagano de potássio em meio ácido. A reação segue a equação: 2KMnO + 5H2O2 + 4 H2 SO + 2 KMnSO + 8H2O +5O2 A diferença entre os números de titulação do controle e do experimento corresponde à quantidade de peróxido de hidrogênio destruído sob a ação da catalase. Valores padrão: Método de titulação: catalase número 12-20. Em um µl. O sangue dos homens contém 4,10-5,10 eritrócitos, o sangue das mulheres contém 3,9,10 - 4,7,10 eritrócitos. Método fotométrico: segundo E. Beutler 15,31·10 ± 2,39·10 UI/g de hemeglobina. Observa-se aumento da atividade da catalase em quadros hemolíticos, durante intervenções cirúrgicas realizadas sob anestesia local, em crianças com patologia broncopulmonar, artrite reumatóide e bócio tireotóxico. Observa-se diminuição da atividade da catalase em doenças infecciosas, anemia ferropriva, síndrome de má absorção, carcinoma, em recém-nascidos com síndrome do desconforto respiratório, intoxicação crônica por fósforo, arsênico, chumbo, mercúrio, anestesia geral e prescrição de antibióticos. É aconselhável determinar a atividade da catalase com um estudo simultâneo da atividade de outras enzimas AO, do conteúdo de metemoglobina e de produtos de peroxidação lipídica. O método mais comum para determinar a atividade GR é o método espectrofotométrico. Entre outros métodos, destaca-se a determinação colorimétrica da atividade do GR pela taxa de formação de glutationa reduzida durante a titulação iodométrica ou por reação de cor com nitroprusida. Determinação da atividade GR pelo método Beutler. Princípio do método: o método baseia-se na determinação espectrofotométrica da quantidade de coenzima NADPH consumida durante a reação enzimática, levada em consideração pela variação da absorção a 340 nm. O comprimento de onda de 340 nm corresponde à absorção máxima da coenzima reduzida. Durante a reação, à medida que o NADPH é oxidado, a densidade óptica da amostra de incubação diminui. A atividade do GR está significativamente aumentada no soro de pacientes com sarcoma, em pacientes com carcinoma de mama e em outras doenças neoplásicas. Um aumento no GH é observado na hepatite, icterícia obstrutiva e, menos comumente, na cirrose. Valores elevados de atividade podem ser determinados durante o infarto agudo do miocárdio. Via de regra, a anemia megaloblástica é acompanhada por um aumento no nível de atividade do GH. Glutationa peroxidase (GP). Catalisa a reação de oxidação da glutationa G-SH com peróxido de hidrogênio (1ª reação) ou hidroperóxidos ROOH (2ª reação), formados a partir do LPO. A contribuição do HP para a neutralização do peróxido de hidrogênio é mais significativa em comparação à catalase. Tem maior afinidade pelo peróxido de hidrogênio e o decompõe mesmo em baixas concentrações. O sítio ativo da enzima contém selênio. Existem 4 gramas de átomo de selênio por 1 mol de enzima. Determinação da atividade GP pelo método de E. Beutler. Princípio do método: o método baseia-se na determinação da taxa de formação de glutationa oxidada, cujo conteúdo é determinado na reação acoplada da glutationa redutase pelo grau de oxidação do NADPH a 340 nm. G - S - S - G + NADPH→2G - SH + NADP O hidroperóxido de T-butila é usado como hidroperóxido. Valores padrão: Segundo Beutler 30,8±4,73 UI/g de hemoglobina (na coleta de sangue com EDTA) e 34,2±3,84 UI/g de hemoglobina (em sangue heparinizado). Segundo Titz: 19,9±0,31 UI/mol de hemoglobina e 0,89±0,14 SEMANA/eritrocito. Aumento da atividade é observado com deficiência de glicose-6-fosfato desidrogenase , α- talassemia, leucemia linfocítica aguda. Uma diminuição na atividade da GP é observada na anemia por deficiência de ferro, envenenamento por chumbo, anemia falciforme e deficiência de selênio. Ceruloplasmina (cobre oxidase). Glicoproteína contendo cobre α2- fração de globulina. As funções da ciruloplasmina são diversas: é a principal oxidase do plasma sanguíneo, um reagente de fase aguda, e transporta Cu para a síntese de SOD e citocromo oxidase. A ciruloplasmina exibe suas propriedades AO extracelularmente sem a formação de quaisquer radicais. A proteína ceruloplasmina pura tem uma cor azul intensa. O método colorimétrico simples proposto por Ravin é amplamente utilizado na modificação (S.V. Bestuzhevoy e V.G. Kolba). Existem outros métodos de pesquisa: nefelométrico, manométrico e imunológico. Com o método manométrico, a reação ocorre em um aparelho Warburg; a taxa de consumo de oxigênio é medida (μmol de oxigênio consumido em 1 minuto por 1 litro de soro sob certas condições). Observou-se excelente correlação entre os valores obtidos pelo método colorimétrico Revin e pelo método monométrico. O método colorimétrico foi modificado para uso em um analisador automático. Determinação da atividade da ciruloplasmina pelo método Revin modificado. Princípio do método: o método baseia-se na oxidação não enzimática da n-fenilenodiamina com ceruloplasmina. A reação é interrompida pela adição de fluoreto de sódio. A densidade óptica dos produtos coloridos resultantes é usada para avaliar a concentração de ceruloplasmina. O nível de ceruloplasmina no soro aumenta em diversas doenças infecciosas, em processos inflamatórios agudos e crônicos acompanhados de alterações destrutivas e necróticas nos tecidos, no crescimento maligno e na esquizofrenia. A atividade antioxidante (AOA) é um complexo de reações enzimáticas e não enzimáticas de ligação e decomposição de produtos intermediários de peroxidação que inibem a oxidação radical livre de lipídios. Na maioria das vezes é determinado por quimiluminescência em sistemas modelo. O valor normal do soro sanguíneo é de 60-75%. Um alto nível de AOA garante resistência aos danos do peróxido nas membranas celulares e um baixo nível de peroxidação lipídica. Um baixo nível de AOA promove aumento da peroxidação lipídica, inibição da proliferação e processos de regeneração. O significado clínico da determinação da AOA é que alguns processos patológicos se desenvolvem no contexto do aumento da AOA, enquanto outros se desenvolvem no contexto da diminuição da AOA e, portanto, requerem correção multidirecional. . Conclusão Assim, a base para a manutenção da homeostase dos radicais livres é o equilíbrio entre os processos pró-oxidantes e antioxidantes que mantêm a peroxidação dentro de limites que não são apenas compatíveis com a vida, mas também benéficos para ela. A violação deste equilíbrio é o ponto de partida para o início da “patologia dos radicais livres”. A quebra da proteção antioxidante é caracterizada pelo desenvolvimento de danos causados pelos radicais livres em vários componentes das células e tecidos. Todos os componentes celulares são suscetíveis à peroxidação em um grau ou outro, mas esse processo é mais pronunciado nas estruturas lipídicas (fosfolipídicas), principalmente na bicamada lipídica das membranas. A violação do equilíbrio pró e antioxidante, acompanhada por um aumento na concentração de produtos de peroxidação lipídica nos tecidos e fluidos do corpo, tem sido observada em uma ampla variedade de doenças: doenças inflamatórias, cardiovasculares, oncológicas, infecciosas, queimaduras e radiação , com diversos efeitos tóxicos, bem como com o envelhecimento do organismo. Tudo isso permite aos pesquisadores considerar a ativação do LPO como um componente universal da resposta inespecífica do corpo a influências extremas, ou seja, como parte da resposta ao estresse. Do exposto conclui-se que o mecanismo de manutenção do equilíbrio pró-oxidante-antioxidante é bastante complexo. Por um lado, atuam fatores e substratos oxidantes e pró-oxidantes: oxigênio molecular, OH ˙, hidroperóxidos, peróxidos orgânicos, epóxidos, substratos facilmente oxidáveis (lipídios), enzimas oxidativas e íons metálicos livres com valência variável, neurotransmissores (catecolaminas). Por outro lado, componentes antioxidantes; enzimas (SOD, GPO, glutationa transferase, catalase), hormônios (esteróide e tireoide), bioaminas (serotonina, histamina), antioxidantes solúveis em gordura - componentes de membrana (tocoferóis, ubiquinonas, retinóides, carotenóides, compostos fenólicos); antioxidantes solúveis em água (compostos tiol, ascorbato, fenóis solúveis em água); Íons de selênio - livres e como parte de enzimas antioxidantes. Para diagnosticar laboratorialmente a gravidade do estresse oxidativo e determinar as indicações para o uso de medicamentos com propriedades antioxidantes, é aconselhável avaliar o estado do sistema antioxidante, bem como a intensidade das reações de radicais livres (processos LPO) no corpo do paciente . Como resultado da análise da literatura, podemos concluir que para avaliar o estado da intensidade da peroxidação lipídica no corpo humano, é necessária a utilização de abordagens integrativas, a utilização de apenas um dos testes acima não; fornecer informações completas. Além disso, o diagnóstico laboratorial da intensidade da peroxidação lipídica é necessário para avaliar a atividade AO de um determinado medicamento, uma vez que com base nos sinais clínicos é impossível avaliar adequadamente o efeito do medicamento AO. Lista de fontes usadas Dubinina, E. E. Estresse oxidativo como reação de adaptação do corpo a condições extremas / E. E. Dubinina // Questões de química médica. - 2001. - Nº 6., volume 47 - P. 561-581. Balabolkin, M. I. O papel do estresse oxidativo na patogênese das complicações vasculares do diabetes / M. I. Balabolkin, E. M. Klebanova // Problemas de endocrinologia. - 2000. - Nº 6. - P. 29-34. Vladimirov, Yu.A. Radicais livres nos principais sistemas / Yu.A. Vladimirov, O.A. Azizova; editado por IA Deev. - M.: VINITI, série biofísica, 1991. - 252 p. Radicais livres. Definição, nomenclatura, classificação [recurso eletrônico] / Acadêmico da Academia Russa de Ciências Médicas, Professor Vladimirov Yu.A. - 2006. - Modo de acesso: #"justify">Vladimirov, Yu.A. Peroxidação lipídica em membranas biológicas / Yu.A. Vladimirov, A.I. Arcakov. - M.: Medicina, 1972. - 252 p. Shanin, Yu.N. Terapia antioxidante na prática clínica / Yu.N. Shanin, V. Yu Shanin, EV Zinoviev. - São Petersburgo, ELBI - São Petersburgo, 2003 - 128 p. Zaichik, Ash. Fisiopatologia geral (com noções básicas de imunologia) / A.Sh. Zaichik, L.P. Churilo. - São Petersburgo, ELBI - São Petersburgo, 2005. - 656 p. Kashulina, A.P. O papel da oxidação do peróxido de radical livre na patologia e métodos de seu estudo / A.P. Kashulina, E.N. consultando. - 1996. - Nº 2. - P. 20-24. Bueverov, A. O. Estresse oxidativo e seu papel na lesão hepática / A. O. Bueverov. // Jornal Russo de Gastroentorologia. - 2002. - Nº 4. - P. 21-25. Khutsishvili, M. B. Processos de radicais livres e seu papel na patogênese de certas doenças / M. B. Khutsishvili, S. I. Rapoport. // Medicina Clínica. - 2002. - Nº 10. - P. 10-16. Tróia, C.M. A regulação negativa da superóxido dismutase de cobre e zinco causa morte apoptótica em células neuronais PC 12 / C.M. Tróia // Proc. Nacional. Acad. Ciência. EUA. - 1994. - Nº 14. - S. 6384 - 6387. Boldyrev, A. A. Introdução à biomembranologia: livro didático. subsídio / Ed. A. A. Boldyreva. - M.: Editora da Universidade Estadual de Moscou, 1997. - 208 p. Danos a componentes de membranas biológicas durante processos patológicos [Recurso eletrônico] / M.: - 2006. - Modo de acesso: #"justify">Radi, R. Peroxidação lipídica de membrana induzida por peroxinitrito: o potencial citotóxico do superóxido e do óxido nítrico / R .Rad //Arco. Bioquímica. Biofísica. - 1991. - Nº 2. - S. 481-487. Danilova, L. A. Coleção de métodos de pesquisa laboratorial / Ed. L. A. Danilova. - São Petersburgo: Peter, 2003. - 736 p. Lifshits, V.M. V. I. Sidelnikova. ″Testes laboratoriais médicos″. Diretório. Segunda edição, corrigida e ampliada / V.M. Lifshits, V.I. Sidelnikova - M.: ″Tríade - X″, 2003. - 312 p. Nikolaev, A. Ya. Química Biológica / Nikolaev A. Ya. - M.: Agência de Informação Médica, 2001. - 496 p.Trabalhos semelhantes a - Diagnóstico laboratorial da intensidade da peroxidação lipídica
Oxidação de peróxido um complexo processo em cadeia de vários estágios de oxidação de oxigênio de substratos lipídicos, principalmente ácidos graxos poliinsaturados, incluindo os estágios de interação de lipídios com compostos de radicais livres e a formação de radicais livres de natureza lipídica. O. p. os fosfolipídios das membranas biológicas desempenham um papel importante na vida dos organismos vivos. O fortalecimento dos processos de O. p. é essencial na etiologia e patogênese de muitas doenças e no desenvolvimento das consequências de diversas influências extremas. A oxidação por peróxido é um caso especial de oxidação em fase líquida de hidrocarbonetos. É um processo típico em cadeia com ramificação pronunciada. O. p. pode incluir estágios de autooxidação não enzimática e reações enzimáticas. As vias enzimáticas e não enzimáticas de produção de oxigênio levam à formação de radicais lipídicos livres em várias etapas principais: iniciação (nucleação da cadeia), continuação da cadeia; ramo de cadeia Os produtos O. p., em particular os peróxidos lipídicos, são utilizados no corpo para a síntese de substâncias biologicamente ativas - prostaglandinas (Prostaglandinas), tromboxanos, hormônios esteróides (Hormônios), etc. A intensidade de O. está diretamente relacionada aos processos de renovação da composição dos fosfolipídios nas membranas biológicas, alterações no conteúdo relativo de lipídios e proteínas e, consequentemente, alterações na estrutura das membranas biológicas e seu funcionamento. Nos organismos vivos, existe uma regulação complexa da intensidade do processo de produção de oxigênio. Normalmente, os processos de formação e consumo de produtos de oxigênio são bem equilibrados, o que determina seu conteúdo relativamente baixo nas células. A taxa de produção de oxigênio nos níveis de iniciação, continuação e terminação da cadeia é amplamente determinada pela organização estrutural dos lipídios na membrana biológica. que afeta os resíduos de ácidos graxos insaturados (ácidos graxos) para oxigênio. Fatores que perturbam o “empacotamento” de lipídios em uma membrana biológica aceleram, e fatores que apoiam a estruturação de lipídios (por exemplo), inibem O. p. Outro componente regulador do sistema O. p. por exemplo, algumas) ou morte (superóxido dismutase) espécies reativas de oxigênio e radicais livres, bem como na decomposição de peróxidos sem formação de radicais livres (catalase, peroxidases). Estas enzimas também podem depender da estruturação da bicamada lipídica da membrana biológica. Em quase todos os estágios da opiáceos, um papel modulador significativo é desempenhado por fatores que regulam os fosfolipídios das membranas biológicas e influenciam a taxa de oxidação, alterando a composição lipídica das membranas. Numerosos compostos de baixo peso molecular que desempenham as funções de iniciadores, catalisadores, inibidores, supressores e sinérgicos deste processo são extremamente importantes na regulação das opacidades. Entre os estabilizadores mais importantes das membranas biológicas está o antioxidante natural (O.p.) E; outros antioxidantes naturais são tiroxina e vitamina K. Os íons metálicos de valência variável, C, D, etc., possuem propriedades de pró-oxidantes (substâncias que aumentam a absorção de oxigênio). Com o desenvolvimento de um processo patológico, o equilíbrio de formação e consumo de peróxidos e outros produtos de oxigênio pode ser perturbado, acumulando-se em tecidos e fluidos biológicos, o que leva a graves distúrbios, principalmente nas membranas biológicas; A consequência da ativação de O. p pode ser uma mudança nas propriedades físico-químicas das proteínas e lipídios da membrana, uma mudança na atividade das enzimas ligadas à membrana, uma violação da permeabilidade da membrana (incluindo prótons e íons de cálcio), transporte de íons. (por exemplo, inibição da bomba de sódio), reduzindo a estabilidade elétrica da bicamada lipídica das membranas. A ativação de O. p. leva a alterações na estrutura das lipoproteínas séricas e hipercolesterolemia, perturba vários processos do metabolismo celular em quase todos os níveis. Não apenas os peróxidos formados como resultado da opiáceos são tóxicos para o corpo, mas também os produtos da oxidação mais profunda de lipídios, aldeídos e ácidos. Os produtos carbonílicos de O. p. inibem uma série de enzimas, suprimem o DNA, aumentam os capilares, modificam a agregação plaquetária e exibem vários outros efeitos indesejáveis. Os radicais livres reativos que iniciam o oxigênio e surgem durante o processo de oxidação causam as estruturas dos ácidos nucléicos (ácidos nucleicos), principalmente DNA, a destruição de coenzimas de nucleotídeos (Coenzimas), interrupção do funcionamento de enzimas (principalmente enzimas SH) e covalente modificação de várias biomoléculas. A consequência da geração excessiva de radicais livres pode ser alterações patológicas nas propriedades dos vasos sanguíneos. Para a prevenção e tratamento de condições associadas à ativação excessiva de O. p., substâncias que reagem especificamente com certos radicais livres (armadilhas ou interceptores), substâncias específicas que formam compostos complexos com metais de valência variável, bem como diversas formas de ativação sistemas endógenos de defesa antirradical do corpo (por exemplo, gradual à hipóxia ou outros fatores). Em conexão com o importante papel de O. na patogênese de diversas doenças, a determinação dos produtos desse processo (principalmente dienos conjugados, malondialdeído), quimiluminescência espontânea e induzida em material biológico (soro e plasma sanguíneo, glóbulos vermelhos, urina , condensado de ar exalado, etc.) d.) tem um significado diagnóstico e prognóstico cada vez maior. 1. Pequena enciclopédia médica. - M.: Enciclopédia Médica. 1991-96 2. Primeiros socorros. - M.: Grande Enciclopédia Russa. 1994 3. Dicionário Enciclopédico de Termos Médicos. - M.: Enciclopédia Soviética. - 1982-1984.
![]() ; produtos moleculares de terminação de cadeia, produtos moleculares, produtos moleculares, onde RH é o substrato de oxidação (ácido graxo poliinsaturado). As chamadas espécies reativas de oxigênio, principalmente radicais de oxigênio contendo elétrons desemparelhados, desempenham um papel decisivo no início da opacificação. Como resultado da redução de um elétron do oxigênio molecular O 2 nas células, forma-se um radical ânion superóxido, que ocorre na cadeia de transferência de elétrons das mitocôndrias, cloroplastos, em reações catalisadas por certas enzimas oxidativas, durante a autooxidação de monoaminas e outros compostos. Durante a reação de dismutação de dois radicais superóxido, forma-se uma molécula de peróxido de hidrogênio H 2 O 2; Outras fontes de peróxido de hidrogênio incluem reações catalisadas por certas oxidases. As células possuem sistemas especiais para neutralizar radicais tóxicos de oxigênio, em particular sistemas enzimáticos: superóxido dismutase, que catalisa a conversão de superóxido em peróxido de hidrogênio, e peroxidases, que catalisam reações nas quais o peróxido de hidrogênio é reduzido a água. Os radicais de oxigênio mais reativos e, portanto, mais perigosos incluem o radical hidroxila OH - um dos principais fatores prejudiciais quando exposto à radiação ionizante (radiação ionizante). Uma parte significativa dos radicais OH nos organismos vivos é gerada como resultado de reações de radicais peróxido de hidrogênio e superóxido com quantidades catalíticas de metais de valência variável, principalmente com íons e cobre. H 2 O 2 relativamente pouco ativo e de longa duração pode servir como fonte do radical OH, que interage com quase todas as classes de biomoléculas na presença de vestígios de ferro ou cobre livre. Juntamente com o radical OH, outros radicais livres, por exemplo, o ânion superóxido protonado, bem como o singleto e uma série de outras espécies reativas de oxigênio, podem ser iniciadores diretos de reações reativas ao oxigênio.
; produtos moleculares de terminação de cadeia, produtos moleculares, produtos moleculares, onde RH é o substrato de oxidação (ácido graxo poliinsaturado). As chamadas espécies reativas de oxigênio, principalmente radicais de oxigênio contendo elétrons desemparelhados, desempenham um papel decisivo no início da opacificação. Como resultado da redução de um elétron do oxigênio molecular O 2 nas células, forma-se um radical ânion superóxido, que ocorre na cadeia de transferência de elétrons das mitocôndrias, cloroplastos, em reações catalisadas por certas enzimas oxidativas, durante a autooxidação de monoaminas e outros compostos. Durante a reação de dismutação de dois radicais superóxido, forma-se uma molécula de peróxido de hidrogênio H 2 O 2; Outras fontes de peróxido de hidrogênio incluem reações catalisadas por certas oxidases. As células possuem sistemas especiais para neutralizar radicais tóxicos de oxigênio, em particular sistemas enzimáticos: superóxido dismutase, que catalisa a conversão de superóxido em peróxido de hidrogênio, e peroxidases, que catalisam reações nas quais o peróxido de hidrogênio é reduzido a água. Os radicais de oxigênio mais reativos e, portanto, mais perigosos incluem o radical hidroxila OH - um dos principais fatores prejudiciais quando exposto à radiação ionizante (radiação ionizante). Uma parte significativa dos radicais OH nos organismos vivos é gerada como resultado de reações de radicais peróxido de hidrogênio e superóxido com quantidades catalíticas de metais de valência variável, principalmente com íons e cobre. H 2 O 2 relativamente pouco ativo e de longa duração pode servir como fonte do radical OH, que interage com quase todas as classes de biomoléculas na presença de vestígios de ferro ou cobre livre. Juntamente com o radical OH, outros radicais livres, por exemplo, o ânion superóxido protonado, bem como o singleto e uma série de outras espécies reativas de oxigênio, podem ser iniciadores diretos de reações reativas ao oxigênio.
Veja o que é “oxidação de peróxido” em outros dicionários:
peroxidação lipídica- O processo de interação dos lipídios (suas seções insaturadas), que fazem parte das membranas celulares, com agentes oxidantes (ânion O2, radical HO, etc.) formados sob a influência da radiação ionizante e nos processos metabólicos de certas substâncias; ... Guia do Tradutor Técnico
Mecanismo de LPO A peroxidação lipídica (LPO) é a degradação oxidativa de lipídios, ocorrendo principalmente sob a influência de radicais livres. Um dos principais... Wikipedia
Peroxidação lipídica Peroxidação lipídica. O processo de interação dos lipídios (suas seções insaturadas), que fazem parte das membranas celulares, com agentes oxidantes (ânion O2, radical HO, etc.) formados sob a influência de ionizantes... ... Biologia molecular e genética. Dicionário.
O conjunto de processos de digestão e absorção de gorduras neutras (triglicerídeos) e seus produtos de degradação no trato gastrointestinal, metabolismo intermediário de gorduras e ácidos graxos e remoção de gorduras, bem como seus produtos metabólicos do corpo.... . .. Enciclopédia Médica - conjunto de processos de absorção, distribuição, assimilação e excreção de substâncias minerais encontradas no corpo principalmente na forma de compostos inorgânicos. Os minerais desempenham um papel importante na manutenção do ácido-base... ... Enciclopédia médica
Mecanismo POL. A peroxidação lipídica (LPO) é a degradação oxidativa dos lipídios, ocorrendo principalmente sob a influência de radicais livres. Uma das principais consequências da radiação. Um dos produtos desse processo é o malondialdeído. Literatura Yu ... Wikipedia
Árvores jovens em um jardim botânico... Wikipedia
- (sinônimo: gorduras neutras, triglicerídeos) ésteres de álcool triatômico glicerol e ácidos graxos superiores ou médios, principal componente das gorduras animais e óleos vegetais, estão presentes em todos os tecidos animais e vegetais, na nutrição... ... Enciclopédia médica
O efeito caotrópico do excesso de ácidos graxos e lisofosfatídeos apoia a ativação peroxidação lipídica (LPO), iniciado pelo acúmulo de espécies reativas de oxigênio (ROS) em uma célula hipóxica. A geração deste último está associada a danos às mitocôndrias dependentes de Ca 2+ e à formação de um excesso de doadores de elétrons - cofatores reduzidos.
Formação de espécies reativas (tóxicas) de oxigênio (em um estado não excitado, o oxigênio não é tóxico) está associada às peculiaridades de sua estrutura molecular: O 2 contém dois elétrons desemparelhados com spins paralelos, que não podem formar um par termodinamicamente estável e estão localizados em orbitais diferentes. Cada um desses orbitais pode aceitar mais um elétron. Assim, a redução completa de uma molécula de oxigênio ocorre como resultado de quatro transferências de um elétron:
E - e - e - e - , H +
O 2 O 2 - H 2 O 2 `OH + H 2 O 2H 2 O
Superóxido formado durante a redução incompleta de moléculas de oxigênio (O 2 -), peróxido (H2O2) e radical hidroxila (`LIGADO) – espécies que reagem ao oxigênio , são agentes oxidantes que representam um sério perigo para muitos componentes estruturais da célula (Avdeeva L.V., Pavlova N.A., Rubtsova G.V., 2005). O radical hidroxila (OH) é especialmente ativo, interagindo com a maioria das moléculas orgânicas. Ele tira um elétron deles e, assim, inicia reações em cadeia de oxidação.
A principal rota de formação de ROS na maioria das células - vazamento de elétrons de sua cadeia de transmissão (cadeia respiratória) e a interação direta desses elétrons com o oxigênio (Gubareva L.E., 2005). Como mais duas fontes pode realizar reações envolvendo oxidases , usando oxigênio molecular como aceptor de elétrons e reduzindo-o a H 2 O ou H 2 O 2 e reações envolvendo oxigenases, incluindo um (monooxigenase) ou dois (dioxigenase) átomos de oxigênio no produto de reação resultante. Sob condições de deficiência de oxigênio nos tecidos, ou seja, numa situação em que a “demanda” (cofatores reduzidos) excede a “oferta” (o número de moléculas de oxigênio), a probabilidade de aumento da formação de ERO aumenta acentuadamente. As reações de radicais livres que iniciam levam a danos nas estruturas celulares e subcelulares, incluindo mitocôndrias, DNA e moléculas de proteínas. E embora a contribuição das ERO para o desenvolvimento da necrobiose hipóxica (em oposição à síndrome de reperfusão) não seja considerada o mecanismo dominante por todos os autores (Zaichik A.Sh., Churilov L.P., 1999), no entanto, sua participação na ativação de os processos de radicais livres na célula, incluindo LPO, são decisivos.
Ressalta-se que a LPO, sendo uma reação em cadeia de autodesenvolvimento, ocorre constantemente na célula, desempenhando o papel de elo necessário em sua atividade vital e nas reações adaptativas. Devido à peroxidação, grupos hidroperóxidos polares (hidroperóxidos lipídicos), que têm efeito detergente, aparecem na molécula fosfolipídica das membranas celulares contendo um ácido graxo na segunda posição. O aparecimento de tais grupos aumenta a mobilidade das cadeias polipeptídicas, ou seja, facilita mudanças conformacionais nas moléculas de proteínas, que são acompanhadas por um aumento na atividade das enzimas ligadas à membrana, que incluem essencialmente todos os sistemas enzimáticos da célula. E apenas a ativação excessiva do LPO, afetando mais de 3-5% dos fosfolipídios da membrana, o transforma de um mecanismo regulador em um elo na patogênese de seus danos durante a morte celular (Yu.A. Vladimirov, 1987; 2000).
Como resultado da ativação do LPO iniciada pelas ROS, e principalmente pelo radical hidroxila (OH), ocorre a formação de novos radicais secundários: lipídico (L), alcóxi (LO), peróxido (LOO). Arroz. 28.

Arroz. 28. Peroxidação lipídica e formação de radicais secundários
(Yu.A. Vladimirov, 2001)
A atividade química desses radicais orgânicos secundários é inferior à do radical hidroxila (OH), mas estão ativamente envolvidos na reação em cadeia da peroxidação lipídica, mantendo e agravando os danos à bicamada lipídica das membranas celulares.
Os efeitos modificadores do LPO nos fosfolipídios determinam a cadeia de eventos adicionais (Arkhipenko Yu.V. et al., 1983; Meerson F.Z., 1989; Vladimirov Yu.A., 2001). Em primeiro lugar, nas moléculas de fosfolipídios contendo um ácido graxo na segunda posição, aparece um grupo hidroperóxido polar (Fig. 29).
Nesse caso, o acúmulo de hidroperóxidos lipídicos é acompanhado por uma diminuição na quantidade de lipídios insaturados. No ativação moderada LPO, conforme observado acima, o aparecimento no microambiente de proteínas integrais de produtos polares de LPO, que possuem efeito detergente, provoca um aumento na mobilidade da cadeia polipeptídica, que geralmente é acompanhado por um aumento na atividade catalítica das enzimas . No ativação excessiva A LPO é de fundamental importância para reduzir a quantidade de fosfolipídios insaturados.

Arroz. 29. Formação de hidroperóxido fosfolipídico, etapa inicial do processo de peroxidação lipídica
(FZ Meerson, 1984).
· Uma diminuição significativa no conteúdo de fosfolipídios insaturados na membrana sob a influência do LPO aumenta a rigidez (microviscosidade) de sua bicamada lipídica, que é acompanhada por uma diminuição na mobilidade conformacional das cadeias polipeptídicas de proteínas incorporadas na membrana (o efeito de “congelamento”). Como essa mobilidade é necessária para o funcionamento normal de enzimas, receptores e formadores de canais, sua resposta funcional é inibida(Fig. 30) .

Arroz. trinta Mudanças na atividade da Ca-ATPase nas membranas sarcoplasmáticas
retículo como resultado da modificação do ambiente lipídico desta enzima
Processo de LPO(FZ Meerson, 1984)
A - estado inicial; B - ativação moderada de Ca-ATPase; B - inibição da Ca-ATPase.
· Os fosfolipídios oxidados durante a ativação do LPO sofrem difusão lateral ao longo da membrana e formam associados (aglomerados) fixados pela interação dos fosfolipídios entre si e com moléculas de água. Essas áreas da membrana tornam-se hidrofílicas. Localizados frente a frente em cada uma das monocamadas da bicamada lipídica, tais associados formam canais na membrana, aumentando sua permeabilidade à água, ao cálcio e a outros íons.(Fig. 31).

Arroz. 31. Esquema de formação de aglomerados de peróxido e fragmentação da membrana durante a indução da peroxidação lipídica (F.Z. Meerson, 1984)
O triângulo claro é o grupo hidroperóxido.
· Os produtos de decomposição resultantes de hidroperóxidos fosfolipídicos (malônicos, glutáricos e outros dialdeídos) interagem com grupos amino livres de proteínas de membrana, formando ligações cruzadas intermoleculares e inativando essas proteínas(Fig. 32) . In vivo, esse processo leva à formação dos chamados. As bases de Schiff da lipofuscina usam pigmento.

Arroz. 32. Formação de ligações cruzadas e inibição de enzimas proteicas de membrana como resultado da ativação de LPO(FZ Meerson, 1984)
Este último é uma mistura de lipídios e proteínas interligados por ligações covalentes transversais e desnaturados como resultado da interação com grupos quimicamente ativos (dialdeídos) de produtos de peroxidação lipídica. Esse pigmento é fagocitado, mas não é hidrolisado pelas enzimas lisossomais e, portanto, acumula-se nas células na forma de manchas pigmentares, principalmente na superfície dorsal das palmas das mãos dos idosos.
O hidroperóxido (2), formado como resultado da reação dos fosfolipídios (1) com o oxigênio molecular, se decompõe em um fosfolipídio com uma cadeia de hidrocarbonetos encurtada na segunda posição, semelhante aos lisofosfolipídios (3) e um fragmento curto de hidrocarbonetos - dialdeído (4 ). A interação de uma molécula de dialdeído, de natureza bifuncional, com os grupos amino de duas moléculas de proteína leva simultaneamente à formação de uma ligação cruzada (5).
· Sob a influência do LPO, grupos sulfidrila (-SH) de proteínas de membrana: enzimas, canais iônicos e bombas são oxidados, o que leva à diminuição de sua atividade.
· A formação de produtos de oxidação polar contribui para o aumento da carga superficial negativa da membrana, o que provoca a fixação de polieletrólitos nela. Entre estes últimos estão algumas proteínas e peptídeos que formam poros proteicos - um dos fatores que reduzem a estabilidade elétrica das membranas.
· Um aumento na polaridade da membrana interna da membrana provoca a penetração de água na bicamada lipídica - a chamada. "corrosão de membrana à base de água".
· “Expulsar” alguns dos ácidos graxos poliinsaturados oxidados da membrana leva a uma diminuição na área de sua bicamada lipídica.
Assim, nesta fase de desenvolvimento do dano celular hipóxico, o elo chave na patogênese é a desorganização da bicamada lipídica das membranas, realizada com a participação dos íons cálcio e da tríade lipídica: ativação de lipases e fosfolipases; ação detergente do excesso de ácidos graxos e lisofosfolipídios, e ativação da peroxidação lipídica.
Uma contribuição significativa para esta desorganização também é feita por: estiramento mecânico (osmótico) de membranas e adsorção de polieletrólitos na bicamada lipídica , contribuindo para o aumento da sua porosidade. Em conjunto, essas violações causam uma diminuição na resistência elétrica das membranas e a ocorrência de quebra elétrica da bicamada lipídica pelo seu próprio potencial de membrana(Fig. 33). Este último é considerado um mecanismo terminal para perturbar a função de barreira da membrana (Vladimirov Yu.A., 2001).
Esta etapa da cadeia patogenética de dano celular durante a hipóxia, caracterizada aumentando a perda de funções de barreira e matriz das membranas, determina transição de mudanças reversíveis na célula para irreversíveis.
Os desenvolvimentos subsequentes de eventos estão associados à formação danos às estruturas celulares, levando diretamente à morte celular. É significativo que os mecanismos destes efeitos prejudiciais também estejam intimamente relacionados com o aumento do conteúdo de íons Ca 2+ no citosol.
As consequências patogenéticas do excesso de íons cálcio no estágio final do dano celular hipóxico (estágio de necrobiose) não estão limitadas a ativação de lipases e fosfolipases. Os íons Ca 2+ estão diretamente envolvidos nos efeitos diretos de danos às estruturas celulares e morte celular por apoptose. Esses efeitos incluem:
· Destruição do citoesqueleto, que está associado Ativação dependente de Ca 2+ de calpaínas. Ocorre destruição de algumas proteínas citoplasmáticas (β-actina, fodrina), o que causa deformação celular, limitando a possibilidade de sua interação com o microambiente, bem como a capacidade de perceber sinais regulatórios. A fraqueza do citoesqueleto contribui para a desintegração de alguns complexos supramoleculares na célula, em particular, o descolamento dos ribossomos das membranas do retículo endoplasmático rugoso. Como resultado, o citoplasma fica saturado com moléculas de proteínas que sofrem degradação.
· Danos mecânicos às estruturas celulares, condicional Ativação de Ca 2+ da função contrátil das miofibrilas com uma perda simultânea da capacidade de relaxar. Tal contrações de contratura acompanhado por danos mecânicos às estruturas contráteis da célula.
· Saponificação e efeito detergente endógeno. O acúmulo de ácidos graxos na célula na presença de excesso de íons Ca 2+ (e Na +) leva a formação de sabão – sais de ácidos graxos superiores. Por esta razão, a hidrólise de ligações éster é chamada saponificação . A formação de sabões no citosol aumenta acentuadamente sua atividade detergente, que literalmente dissolve as membranas lipídicas (Zaichik A.Sh., Churilov L.P., 1999). Os sabonetes, destruindo as membranas das organelas, atacam a célula com hidrolases, radicais ativos e outros metabólitos, que até aquele momento estavam isolados em vários compartimentos da célula. Este efeito endógeno é crítico na formação do estágio final da morte celular.
· Juntamente com a participação na necrobiose, os íons de cálcio estão envolvidos na implementação mecanismos de morte celular apoptótica. Entre os mais recentes: aumento da atividade de endonucleases e calpaínas dependentes de Ca 2+. Tal ativação representa uma ameaça para a célula, iniciando sua morte apoptótica ou devido à fragmentação do DNA ( endonucleases ), ou como resultado da proteólise de proteínas antiapoptóticas (bcl-2) calpaínas . A apoptose pode ser promovida por degradação induzida por calpaína da proteína quinase C (PKC), percebendo principalmente efeitos antiapoptóticos e aumentando a resistência celular a produtos metabólicos tóxicos.
· Além disso, excesso de íons Ca 2+ eu mesmo promove a formação de produtos tóxicos, que podem, nomeadamente, ser moléculas óxido nítrico em altas concentrações, criado pela ativação de Ca 2+ da NO sintase indutível. Este efeito se manifesta mais claramente com os chamados. morte de neurônios de glutamato ocorrendo durante a hipóxia (isquemia cerebral). O início dos eventos, neste caso, está associado à deficiência de energia nos neurônios, à liberação de íons potássio, à despolarização da membrana e ao aumento do pool intracelular de Ca 2+ como resultado da abertura prolongada de canais de cálcio dependentes de voltagem (Fig. 34 ).
Arroz. 34. O mecanismo de desenvolvimento da morte de neurônios por glutamato durante a hipóxia
A consequência do excesso de íons cálcio no citoplasma é o aumento da liberação do neurotransmissor (glutamato) pelos neurônios glutamatérgicos na fenda sináptica. A percepção desse sinal pelos neurônios pós-sinápticos é realizada por meio de receptores NMDA (o subtipo mais estudado de receptores de glutamato com alta afinidade pelo aminoácido sintético N-metil-D-aspartato), cuja sensibilidade ao transmissor em condições hipóxicas aumenta significativamente (Kryzhanovsky G.N., 1997). O resultado do “bombardeio de glutamato” (Akmaev I.G., 1996; Akmaev I.G., Grinevich V.V., 2001) do neurônio pós-sináptico é a abertura de canais iônicos nele, levando a um aumento no fluxo de cálcio para a célula e ativação de neurônios NO sintase (NOS). O óxido nítrico produzido sob sua influência, de pequeno tamanho e natureza lipofílica da molécula, difunde-se no espaço extracelular e entra através das membranas nas células próximas (neurônios), exercendo sobre elas um efeito tóxico. A base deste efeito tóxico é a deficiência energética das células. O mecanismo para a formação de tal deficiência está associado à capacidade do NO de causar S- nitrosilação de proteínas celulares contendo ferro(ciclo aconitase TCA, complexos I-III da cadeia de transporte de elétrons em MTX) e sua inativação. Além disso, sob a influência do NO, ribosilação E nitrosilaçãogliceraldeído-3-fosfato desidrogenase, causando inibição da glicólise. Finalmente, quando o NO interage com outro radical - O 2 - forma ânion peroxinitrito (ONOO -), causando inibição irreversível de proteínas contendo ferro.
Devido à formação de ONOO, é possível ativar o mecanismo apoptótico de morte celular através da implementação da seguinte cascata:
Uma característica da morte dos neurônios do glutamato é a ausência de morte dos próprios neurônios produtores de NO, que são protegidos dos efeitos tóxicos do NO. O mecanismo dessa proteção está associado à ativação da superóxido dismutase (SOD) e (ou) à transição do NO para a forma oxidada (NO +). Na verdade, existe uma analogia direta com os macrófagos, que, embora produzam NO, apresentam resistência a ele.
Assim, a morte celular durante a hipóxia é um desdobramento natural de uma cadeia de eventos, incluindo a formação de deficiência energética, inibição das principais vias metabólicas, ativação da tríade lipídica e subsequente dano irreversível às estruturas celulares. O elo central na patogênese desses eventos é o aumento da concentração intracelular de íons cálcio, e o principal alvo são as membranas celulares e, principalmente, as mitocôndrias.
A sequência de alterações consideradas durante a hipóxia (anóxia) é a mesma para uma ampla variedade de tecidos. Isto é evidenciado por experimentos com cortes de tecido, células isoladas e organelas isoladas (Vladimirov Yu.A., 2001). Arroz. 35.
A diferença está apenas na velocidade desses processos, que é 2 a 3 vezes maior na temperatura do corpo humano. Além disso, essa velocidade é diferente para diferentes tecidos e esses processos ocorrem em maior velocidade no tecido cerebral, em menor velocidade no fígado e em velocidade ainda menor no tecido muscular.
Arroz. 35. Sequência de distúrbios nas células do fígado durante anóxia
de acordo com Yu.A. Vladimirov, 2001
XIV. HIPERÓXIA
Hiperóxia – aumento do fornecimento de oxigênio ao corpo . Ao contrário da hipóxia, a hiperóxia é sempre exógena e praticamente nunca ocorre em condições naturais. A este respeito, os mecanismos adaptativos a este estado são eficazes apenas sob condições de carga de oxigênio relativamente baixa, determinada pela magnitude da pressão parcial de oxigênio e pela duração de sua ação. Um exemplo dessa dependência é a curva de períodos seguros para respirar oxigênio em humanos (Fig. 36).

Arroz. 36. O limite da ação do oxigênio em humanos(de acordo com Hartmann, 1966).
Citado por A.G. Zhironkin (1979).
O eixo x é a duração da respiração de oxigênio, horas; ao longo da ordenada - pressão parcial de oxigênio, atm.
Como pode ser visto na figura, zona assim chamado "ação fisiológica do oxigênio" dura mais em valores baixos de sua pressão parcial (cerca de 0,5 atm.), quando as reações adaptativas de proteção são capazes de garantir a preservação da tensão normal de oxigênio nos tecidos. Estas reações baseiam-se em mecanismos que visam limitar o fornecimento e transporte de oxigênio. Isto destina-se, em particular, a reação primária do sistema respiratório externo, na forma de diminuição da ventilação pulmonar e do volume respiratório minuto.
Essas alterações são consequência da cessação dos impulsos naturais normais dos quimiorreceptores arteriais sob condições de aumento do suprimento de oxigênio. Ao mesmo tempo, limitar a ventilação não só reduz o fornecimento de oxigênio ao corpo, mas também leva ao desenvolvimento de hipercapnia. Este último determina a segunda fase da reação do sistema respiratório, caracterizada pelo aumento da ventilação visando reduzir a PaCO 2 e eliminar a acidose gasosa. O mais importante mudança no sistema circulatório na hiperóxia, ocorre um estreitamento natural dos pequenos vasos sanguíneos, acompanhado por um aumento da resistência periférica, uma desaceleração do fluxo sanguíneo geral e local e um aumento da pressão diastólica. Outra manifestação da reação desse sistema é a bradicardia, registrada antes do aparecimento de sinais de envenenamento por oxigênio. Mudanças no sistema sanguíneo em resposta à hiperóxia, manifestam-se no período inicial como eritropenia transitória e diminuição dos níveis de hemoglobina, que é causada pelo movimento do fluido tecidual para o sangue e pela deposição de glóbulos vermelhos (Zhironkin A.G., 1979).
À medida que a pressão parcial do oxigênio na mistura gasosa inalada aumenta, seu efeito tóxico vem à tona, uma vez que o efeito protetor das reações adaptativas é minimizado. Nesta zona, o oxigênio já desempenha o papel de fator que não fornece, mas inibe os processos oxidativos nos tecidos. Quanto aos mecanismos do efeito tóxico em si, hoje o ponto de vista mais aceito é o de R. Gershman (1964), que relaciona esse mecanismo com a formação de espécies reativas de oxigênio e com a ativação da oxidação de radicais livres.
Em condições de supersaturação dos tecidos com oxigênio, ou seja, numa situação em que a “oferta” (excesso de oxigênio) excede a “demanda” (a quantidade de cofatores reduzidos sujeitos à oxidação), a probabilidade de aumento da formação de ERO aumenta. Conseqüentemente, a oxidação dos radicais livres aumenta, acompanhada de danos às estruturas celulares e subcelulares e, acima de tudo, às mitocôndrias.
É óbvio que a desorganização e os danos às mitocôndrias serão acompanhados pela ruptura da cadeia de transporte de electrões e pela fosforilação oxidativa. Aqueles. distúrbios que definem a essência do conceito de “hipóxia”. Respectivamente, este estado é chamado hipóxia hiperóxica.
Danos às estruturas celulares e subcelulares durante a ativação de processos de radicais livres levam ao desenvolvimento de numerosos distúrbios nas funções específicas de vários órgãos e sistemas. Assim, a inibição de enzimas cerebrais reduz a produção de ácido γ-aminobutírico, o mediador inibitório mais importante, que serve como um dos mecanismos de desenvolvimento na hiperóxia. síndrome convulsiva de origem cortical. A violação da produção de surfactante pelo epitélio pulmonar provoca uma diminuição acentuada nas reservas compensatórias do sistema respiratório externo, aumentando tensão superficial dos alvéolos e contribuem para o aparecimento de microatelectasias. Em casos graves, a interrupção da produção de surfactante pode ser acompanhada por edema pulmonar. Em algumas crianças no primeiro ano de vida, respirar oxigênio puro leva ao desenvolvimento dificuldade respiratória – displasia broncopulmonar(Malyarenko Yu.E., Pyatin V.F., 1998) . A ativação da oxidação dos radicais livres durante a hiperoxigenação está subjacente à formação defeitos visuais em crianças pequenas devido à maturação prejudicada dos fotorreceptores.
Juntamente com as ERO, o efeito tóxico do oxigênio também é mediado pela tensão excessiva em algumas reações protetoras e adaptativas. Tais reações, em particular, incluem vasoespasmo prolongado (reação à hiperóxia). Em bebês prematuros, promove o desenvolvimento fibroplasia retrolental(formação de tecido fibroso atrás do cristalino), levando à cegueira. Um espasmo semelhante dos vasos sanguíneos nos pulmões causa hipertensão pulmonar, distúrbios da microcirculação e danos ao epitélio pulmonar - distúrbios que predispõem ao desenvolvimento de inflamação.
Estas circunstâncias obrigam-nos a limitar o uso de oxigênio para fins terapêuticos, em que a PO 2 não deve ultrapassar 380 mm Hg. Arte. (Berezovsky V.A., 1975).
O tecido cerebral fetal é particularmente sensível aos efeitos tóxicos do excesso de oxigênio., que é caracterizado por uma tensão de oxigênio significativamente menor do que as estruturas cerebrais de um organismo maduro . “Este facto não resulta de imperfeições nos processos de fornecimento de oxigénio ao corpo no período pré-natal, mas, pelo contrário, reflecte o equilíbrio desses processos, garantindo, por um lado, uma oxigenação adequada do cérebro, e por outro, protegendo-o do excesso de fluxo de O 2 "(Raguzin A.V., 1990). Foi experimentalmente estabelecido que a tensão de oxigênio do tecido cerebral fetal é um parâmetro relativamente estável de homeostase no organismo em desenvolvimento intrauterino, que muda pouco mesmo com mudanças significativas no regime de oxigênio de animais prenhes . Essa constância da PO 2 do tecido cerebral fetal com alterações na PaO 2 (de 50 para 370 mm Hg) do corpo materno é determinada por mecanismos localizados principalmente na região útero-placentária, mas não por reações sistêmicas da respiração e da circulação sanguínea. Por nascimento a formação de mecanismos para estabilizar a homeostase do oxigênio no cérebro não está completa, o que provoca um aumento mais significativo (do que em adultos) da PO 2 das estruturas cerebrais dos recém-nascidos durante a inalação de oxigênio puro. Tal aumento na PO 2 é acompanhado pela ativação da oxidação dos radicais livres no tecido cerebral e pelo desenvolvimento de mudanças qualitativas negativas nos parâmetros dos reflexos defensivos condicionados na idade adulta (Raguzin A.V., 1990). Em conexão com esta situação, uma abordagem para corrigir a hipóxia grave em recém-nascidos é fundamentada usando misturas de gases com conteúdo reduzido em vez de oxigênio puro para inalação.
Forma convulsiva de envenenamento por oxigênio ocorre na intoxicação aguda por oxigênio e é conhecida desde o final do século XIX como Sinal de Baer, descoberto e descrito pela primeira vez por este autor. As convulsões geralmente ocorrem ao respirar oxigênio sob pressão superior a 3-4 atm. e são muito semelhantes em seu curso às crises epilépticas.
Clinicamente, existem três etapas deste processo (Chereshnev V.A., Yushkov B.G., 2001):
Estágio I – aumento da respiração e da frequência cardíaca, aumento da pressão arterial, pupilas dilatadas, aumento da atividade com espasmos musculares ocasionais.
O estágio II é o estágio das crises semelhantes às crises epilépticas com manifestações clônicas e tônicas.
Estágio III – terminal – enfraquecimento das convulsões com dificuldade respiratória, que progride para respirações individuais. A morte ocorre por paralisia do centro respiratório.
Os produtos deste processo incluem malondialdeído e 4-hidroxinonenal.
As reações de oxidação biológica são acompanhadas pela formação de radicais livres, partículas com um elétron desemparelhado na órbita externa. Isso causa a alta atividade química desses radicais. Por exemplo, eles reagem com ácidos graxos insaturados nas membranas, perturbando sua estrutura. Os antioxidantes previnem a oxidação dos radicais livres.
Através da etapa de derivados peróxidos de ácidos graxos insaturados, é realizada a biossíntese de prostaglandinas e leucotrienos, e os tromboxanos, que têm um efeito poderoso nas propriedades de agregação adesiva das células sanguíneas e da microcirculação, são eles próprios hidroperóxidos. A formação de hidroperóxidos de colesterol é um dos elos da síntese de alguns hormônios esteróides, em especial a progesterona.
Literatura
- Vladimirov Yu.A., Archakov A.I. Peroxidação lipídica em membranas biológicas. - M.: Nauka, 1972. - 252 p.
- Baraboy VA, Orel VE, Karnaukh IM. Peroxidação e radiação. - K.: Naukova Dumka, 1991.
- Kovshevny V.V.- oxidação de radicais livres
Notas
Fundação Wikimedia. 2010.
Veja o que é “Peroxidação lipídica” em outros dicionários:
peroxidação lipídica- O processo de interação dos lipídios (suas seções insaturadas), que fazem parte das membranas celulares, com agentes oxidantes (ânion O2, radical HO, etc.) formados sob a influência da radiação ionizante e nos processos metabólicos de certas substâncias; ... Guia do Tradutor Técnico
Peroxidação lipídica Peroxidação lipídica. O processo de interação dos lipídios (suas seções insaturadas), que fazem parte das membranas celulares, com agentes oxidantes (ânion O2, radical HO, etc.) formados sob a influência de ionizantes... ... Biologia molecular e genética. Dicionário.
Um complexo processo em cadeia de múltiplos estágios de oxidação de substratos lipídicos, principalmente ácidos graxos poliinsaturados, com oxigênio, incluindo os estágios de interação de lipídios com compostos de radicais livres e a formação de radicais livres lipídicos... Enciclopédia médica
Mecanismo POL. A peroxidação lipídica (LPO) é a degradação oxidativa dos lipídios, ocorrendo principalmente sob a influência de radicais livres. Uma das principais consequências da radiação. Um dos produtos desse processo é o malondialdeído. Literatura Yu ... Wikipedia
Com o diabetes, o corpo desenvolve falta de vitaminas e minerais. Isso se deve a três motivos: restrições alimentares, distúrbios metabólicos e diminuição da absorção de nutrientes. Por sua vez, deficiência de vitaminas e... ... Wikipedia
- (Dibunol) (ver também acetato de tocoferol). 2.6 Di-terc-butil 4 metilfenol. Sinônimos: Butiloxitolueno, Ionol. Pó cristalino branco ou branco com um tom levemente amarelado. Praticamente insolúvel em água, facilmente solúvel em álcool.... ...
DIBUNOL (Dibunolum) (ver também acetato de tocoferol). 2.6 Di-terc-butil 4 metilfenol. Sinônimos: Butiloxitolueno, Ionol. Pó cristalino branco ou branco com um tom levemente amarelado. Praticamente insolúvel em água, facilmente solúvel em álcool... Dicionário de medicamentos
Ácidos graxos, ácidos carboxílicos; No corpo de animais e plantas, os ácidos graxos livres e os contidos nos lipídios desempenham funções energéticas e plásticas. Os ácidos graxos na composição dos fosfolipídios participam da construção do biológico... ... Enciclopédia médica
Átomos ou grupos de átomos quimicamente ligados que possuem valências livres, ou seja, elétrons desemparelhados (não compensados) no orbital externo (valência). A presença de elétrons desemparelhados determina uma reação química elevada... ... Enciclopédia médica
Princípio ativo ›› Aminoácidos para nutrição parenteral+Outros medicamentos [Emulsões gordurosas para nutrição parenteral + Dextrose + Sais minerais] (Aminoácidos para nutrição parenteral+Outros medicamentos)
Postagens relacionadas:
 Exercícios para desenvolver habilidades de escrita em preparação para a parte de “escrita” do Exame Estadual Unificado em inglês. Exercícios de paráfrase na coleção de inglês.
Exercícios para desenvolver habilidades de escrita em preparação para a parte de “escrita” do Exame Estadual Unificado em inglês. Exercícios de paráfrase na coleção de inglês.
 Resumo sobre o tema: V. Hugo "Les Miserables". "Análise artística do herói do romance Os miseráveis, de Hugo
Resumo sobre o tema: V. Hugo "Les Miserables". "Análise artística do herói do romance Os miseráveis, de Hugo
 Guk "Biblioteca Central Republicana de Polotsk" Realize eventos sobre Nevsky na biblioteca
Guk "Biblioteca Central Republicana de Polotsk" Realize eventos sobre Nevsky na biblioteca
 Equação tangente ao gráfico 10
Equação tangente ao gráfico 10
 Sismologia: como os terremotos são previstos
Sismologia: como os terremotos são previstos