Механизмы возбуждения первичночувствующих рецепторов. Чурилова Т.М
1. Павлова об анализаторах. Структура и функции анализаторов. Механизм возникновения возбуждения в рецепторах. Рецепторный и генераторный потенциалы.
Учение об анализаторах было создано. Анализатором считал совокупность нейронов, участвующих в восприятии раздражений, проведении возбуждения, а также анализе его свойств клетками коры большого мозга. Анализатор впервые рассматривался как единая система, включающая рецепторный аппарат (периферический отдел анализатора), афферентные нейроны и проводящие пути (проводниковый отдел) и участки коры больших полушарий мозга, воспринимающие афферентные сигналы (центральный конец анализатора). Опыты с удалением участков коры и исследованием возникающих вслед за этим нарушений условнорефлекторных реакций привели к заключению о наличии в корковом отделе анализатора первичных проекционных зон (ядерных зон) и так называемых рассеянных элементов, анализирующих поступающую информацию вне ядерной зоны коры большого мозга. Еще до появления современных аналитических (в частности, электрофизиологических) методов исследования сделал доступным для объективного экспериментального анализа пространственно-временное взаимодействие нервных процессов на высших, корковых уровнях анализаторных систем.
Анализаторы – сложные чувствительные образования нервной системы, воспринимающие раздражения из окружающей среды и ответственные за формирование ощущений. Различают три части любого анализатора:
Ø Периферический или рецепторный отдел, который осуществляет восприятие энергии раздражителя и трансформацию ее в специфический процесс возбуждения.
Ø Проводниковый отдел, представленный афферентными нервами и подкорковыми центрами, он осуществляет передачу возникшего возбуждения в кору головного мозга.
Ø Центральный или корковый отдел анализатора, представленный соответствующими зонами коры головного мозга, где осуществляется высший анализ и синтез возбуждений и формирование соответствующего ощущения.
Анализаторы выполняют большое количество функций или операций с сигналами. Среди них важнейшие:
I. Обнаружение сигналов.
II. Различение сигналов.
III. Передача и преобразование сигналов.
IV. Кодирование поступающей информации.
V. Детектирование тех или иных признаков сигналов.
VI. Опознание образов.
Классификация рецепторов. В основу классификации рецепторов положено несколько критериев.
Психофизиологический характер ощущения: тепловые, холодовые, болевые и др.
Природа адекватного раздражителя: механо-, термо-, хемо-, фото-, баро-, осмбрецепторы и др.
Среда, в которой рецептор воспринимает раздражитель: экстеро-, интерорецепторы.
Отношение к одной или нескольким модальностям: моно - и полимодальные (мономодальные преобразуют в нервный импульс только один вид раздражителя - световой, температурный и т. д., полимодальные могут несколько раздражителей преобразовать в нервный импульс - механический и температурный, механический и химический и т. д.).
Способность воспринимать раздражитель, находящийся на расстоянии от рецептора или при непосредственном контакте с ним: контактные и дистантные.
Уровень чувствительности (порог раздражения): низкопороговые (механорецепторы) и высокопороговые (ноцицепторы).
Скорость адаптации: быстроадаптирующиеся, (тактильные), медленноадаптирующиеся (болевые) и неадаптирующиеся (вестибулярные рецепторы и проприорецепторы).
Отношение к различным моментам действия раздражителя: при включении раздражителя, при его выключении, на протяжении всего времени действия раздражителя.
Морфофункциональная организация и механизм возникновения возбуждения: первичночувствующие и вторичночувствующие.
В первичночувствующих рецепторах стимул действует на воспринимающий субстрат, заложенный в самом сенсорном нейроне, который при этом возбуждается непосредственно (первично) раздражителем. К первичночувствующим рецепторам относятся: обонятельные, тактильные рецепторы и мышечные веретена .
К вторичночувствующим относятся те рецепторы, у которых между действующим стимулом и сенсорным нейроном располагаются дополнительные рецептирующие клетки, при этом сенсорный нейрон возбуждается не непосредственно стимулом, а опосредовано (вторично) - потенциалом рецептирующей клетки. К вторичночувствующим рецепторам относятся: рецепторы слуха, зрения, вкуса, вестибулярные рецепторы.
Механизм возникновения возбуждения у этих рецепторов различен. В первичночувствующем рецепторе транформация энергии раздражителя и возникновение импульсной активности идет в самом сенсорном нейроне. У вторичночувствующих рецепторов между сенсорным нейроном и стимулом расположена рецептирующая клетка, в которой под влиянием раздражителя идут процессы трансформации энергии раздражителя в процесс возбуждения. Но в этой клетке не возникает импульсной активности. Рецепторные клетки синапсами соединены с сенсорными нейронами. Под влиянием потенциала рецептирирующей клетки выделяется медиатор, который возбуждает нервное окончание сенсорного нейрона и вызывает в нем появление локального ответа - постсинаптического потенциала. Он оказывает деполяризующее действие на отходящее нервное волокно, в котором возникает импульсная активность.
Следовательно, у вторичночувствующих рецепторов локальная деполяризация возникает дважды: в рецептирующей клетке и в сенсорном" нейроне. Поэтому принято называть градуальный электрический ответ рецептирующей клетки рецепторным потенциалом, а локальную деполяризацию сенсорного нейрона генераторным потенциалом, имея в виду, что он генерирует в отходящем от рецептора нервном волокне распространяющееся возбуждение. У первичночувствующих рецепторов рецепторный потенциал является и генераторным. Таким образом, рецепторный акт можно изобразить в виде следующей схемы.
Для первичночувствующих рецепторов:
I этап - специфическое взаимодействие раздражителя с мембраной рецептора;
II этап - возникновение рецепторного потенциала в месте взаимодействия раздражителя с рецептором в результате изменения проницаемости мембраны для ионов натрия (или кальция);
III этап - электротоническое распространение рецепторного потенциала к аксону сенсорного нейрона (пассивное распространение рецепторного потенциала вдоль нервного волокна называется электротоническим);
IV этап - генерация потенциала действия;
V этап - проведение потенциала действия по нервному волокну в ортодромном направлении.
Для вторичночувствующих рецепторов:
I-III этапы совпадают с такими же этапами первичночувствующих рецепторов, но протекают они в специализированной рецептирующей клетке и заканчиваются на ее пресинаптической мембране;
IV этап - выделение медиатора пресинаптическими структурами рецептирующей клетки;
V этап - возникновение генераторного потенциала на постсинаптической мембране нервного волокна;
VI этап - электротоническое распространение генераторного потенциала по нервному волокну;
VII этап - генерация потенциала действия электрогенными участками нервного волокна;
VIII этап - проведение потенциала действия по нервному волокну в ортодромном направлении.
2. Физиология зрительного анализатора. Рецепторный аппарат. Фотохимические процессы в сетчатке глаза при действии света.
Зрительный анализатор представляет собой совокупность структур, воспринимающих световую энергию в виде электромагнитного излучения с длиной волны нм и дискретных частиц фотонов, или квантов, и формирующих зрительные ощущения. С помощью глаза воспринимается 80-90% всей информации об окружающем мире.
Благодаря деятельности зрительного анализатора различают освещенность предметов, их цвет, форму, величину, направление передвижения, расстояние, на которое они удалены от глаза и друг от друга. Все это позволяет оценивать пространство, ориентироваться в окружающем мире, выполнять различные виды целенаправленной деятельности.
Наряду с понятием зрительного анализатора существует понятие органа зрения.
Орган зрения - это глаз, включающий три различных в функциональном отношении элемента:
Ø глазное яблоко, в котором расположены световоспринимающий, светопреломляющий и светорегулирующий аппараты;
Ø защитные приспособления, т. е. наружные оболочки глаза (склера и роговица), слезный аппарат, веки, ресницы, брови;
Ø двигательный аппарат, представленный тремя парами глазных мышц (наружная и внутренняя прямые, верхняя и нижняя прямые, верхняя и нижняя косые), которые иннервируются III (глазодвигательный нерв), IV (блоковый нерв) и VI (отводящий нерв) парами черепных нервов.
Рецепторный (периферический) отдел зрительного анализатора (фоторецепторы) подразделяется на палочковые и колбочковые нейросенсорные клетки, наружные сегменты которых имеют соответственно палочковидную ("палочки") и колбочковидную ("колбочки") формы. У человека насчитывается 6-7 млн. колбочек и млн. папочек.
Место выхода зрительного нерва из сетчатки не содержит фоторецепторов и называется слепым пятном. Латерально от слепого пятна в области центральной ямки лежит участок наилучшего видения - желтое пятно, содержащее преимущественно колбочки. К периферии сетчатки число колбочек уменьшается, а число палочек возрастает, и периферия сетчатки содержит одни лишь палочки.
Различия функций колбочек и палочек лежит в основе феномена двойственности зрения. Палочки являются рецепторами, воспринимающими световые лучи в условиях слабой освещенности, т. е. бесцветное, или ахроматическое, зрение. Колбочки же функционируют в условиях яркой освещенности и характеризуются разной чувствительностью к спектральным свойствам света (цветное или хроматическое зрение). Фоторецепторы обладают очень высокой чувствительностью, что обусловлено особенностью строения рецепторов и физико-химических процессов, лежащих в основе восприятия энергии светового стимула. Полагают, что фоторецепторы возбуждаются при действии на них 1 - 2 квантов света.
Палочки и колбочки состоят из двух сегментов - наружного и внутреннего, которые соединяются между собой посредством узкой реснички. Палочки и колбочки ориентированы в сетчатке радиально, а молекулы светочувствительных белков расположены в наружных сегментах таким образом, что около 90% их светочувствительных групп лежат в плоскости дисков, входящих в состав наружных сегментов. Свет оказывает наибольшее возбуждающее действие в том случае, если направление луча совпадает с длинной осью палочки или колбочки, при этом он направлен перпендикулярно дискам их наружных сегментов.
Фотохимические процессы в сетчатке глаза. В рецепторных клетках сетчатки находятся светочувствительные пигменты (сложные белковые вещества) - хромопротеиды, которые обесцвечиваются на свету. В палочках на мембране наружных сегментов содержится родопсин, в колбочках - йодопсин и другие пигменты.
Родопсин и йодопсин состоят из ретиналя (альдегида витамина А1) и гликопротеида (опсина). Имея сходство в фотохимических процессах, они различаются тем, что максимум поглощения находится в различных областях спектра. Палочки, содержащие родопсин, имеют максимум поглощения в области 500 нм. Среди колбочек различают три типа, которые отличаются максимумами в спектрах поглощения: одни имеют максимум в синей части спектра (нм), другие в зеленой (, третьи - в красной (нм) части, что обусловлено наличием трех типов зрительных пигментов. Красный колбочковый пигмент получил название "йодопсин". Ретиналь может находиться в различных пространственных конфигурациях (изомерных формах), но только одна из них - 11-ЦИС-изомер ретиналя выступает в качестве хромофорной группы всех известных зрительных пигментов. Источником ретиналя в организме служат каротиноиды.
Фотохимические процессы в сетчатке протекают весьма экономно. Даже при действии яркого света расщепляется только небольшая часть имеющегося в палочках родопсина (около 0,006%).
В темноте происходит ресинтез пигментов, протекающий с поглощением энергии. Восстановление йодопсина протекает в 530 раз быстрее, чем родопсина. Если в организме снижается содержание витамина А, то процессы ресинтеза родопсина ослабевают, что приводит к нарушению сумеречного зрения, так называемой куриной слепоте. При постоянном и равномерном освещении устанавливается равновесие между скоростью распада и ресинтеза пигментов. Когда количество света, падающего на сетчатку, уменьшается, это динамическое равновесие нарушается и сдвигается в сторону более высоких концентраций пигмента. Этот фотохимический феномен лежит в основе темновой адаптации.
Особое значение в фотохимических процессах имеет пигментный слой сетчатки, который образован эпителием, содержащим фусцин. Этот пигмент поглощает свет, препятствуя отражению и рассеиванию его, что обусловливает четкость зрительного восприятия. Отростки пигментных клеток окружают светочувствительные членики палочек и колбочек, принимая участие в обмене веществ фоторецепторов и в синтезе зрительных пигментов.
Вследствие фотохимических процессов в фоторецепторах глаза при действии света возникает рецепторный потенциал, который представляет собой гиперполяризацию мембраны рецептора. Это отличительная черта зрительных рецепторов, активация других рецепторов выражается в виде деполяризации их мембраны. Амплитуда зрительного рецепторного потенциала увеличивается при увеличении интенсивности светового стимула. Так, при действии красного цвета, длина волны которого составляет нм, рецепторный потенциал более выражен в фоторецепторах центральной части сетчатки, а синего (нм) - в периферической.
Синаптические окончания фоторецепторов конвергируют на биполярные нейроны сетчатки. При этом фоторецепторы центральной ямки связаны только с одним биполяром. Проводниковый отдел зрительного анализатора начинается от биполярных клеток, затем ганглиозные клетки, затем зрительный нерв, затем зрительная информация поступает на латеральные коленчатые тела таламуса, откуда в составе зрительной лучистости проецируется на первичные зрительные поля.
Первичными зрительными полями коры является поле 16 и поле 17 – это шпорная борозда затылочной доли.
Для человека характерно бинокулярное стереоскопическое зрения, то есть способность различать объем предмета и рассматривать двумя глазами. Характерна световая адаптация, то есть приспособление к определенным условиям освещения.
3. Слуховой анализатор. Звукоулавливающий и звукопроводящий аппарат органа слуха.
С помощью слухового анализатора человек ориентируется в звуковых сигналах окружающей среды, формирует соответствующие поведенческие реакции, например оборонительные или пищедобывательные. Способность восприятия человеком разговорной и вокальной речи, музыкальных произведений делает слуховой анализатор необходимым компонентом средств общения, познания, приспособления.
Адекватным раздражителем для слухового анализатора являются звуки, т. е. колебательные движения частиц упругих тел, распространяющихся в виде волн в самых различных средах, включая воздушную среду, и воспринимающиеся ухом. Звуковые волновые колебания (звуковые волны) характеризуются частотой и амплитудой. Частота звуковых волн определяет высоту звука. Человек различает звуковые волны с частотой от 20 доГц. Звуки, частота которых ниже 20 Гц - инфразвуки и вышеГц (20 кГц) - ультразвуки, человеком не ощущаются. Звуковые волны, имеющие синусоидальные, или гармонические, колебания, называют тоном. Звук, состоящий из не связанных между собой частот, называют шумом. При большой частоте звуковых волн - тон высокий, при малой - низкий.
Второй характеристикой звука, которую различает слуховая сенсорная система, является его сила, зависящая от амплитуды звуковых волн. Сила звука или его интенсивность воспринимаются человеком как громкость. Ощущение громкости нарастает при усилении звука и зависит также от частоты звуковых колебаний, т. е. громкость звучания определяется взаимодействием интенсивности (силы) и высоты (частоты) звука. Единицей измерения громкости звука является бел, в практике обычно используется децибел (дБ), т. е.0,1 бела. Человек различает звуки также по тембру, или "окраске". Тембр звукового сигнала зависит от спектра, т. е. от состава дополнительных частот (обертонов), которые сопровождают основной тон (частоту). По тембру можно различить звуки одинаковой высоты и громкости, на чем основано узнавание людей по голосу. Чувствительность слухового анализатора определяется минимальной силой звука, достаточной для возникновения слухового ощущения. В области звуковых колебаний от 1000 до 3000 в секунду, что соответствует человеческой речи, ухо обладает наибольшей чувствительностью. Эта совокупность частот получила название речевой зоны. В данной области воспринимаются звуки, имеющие давление меньше чем 0,001 бара (1 бар составляет приблизительно одну миллионную часть нормального атмосферного давления). Исходя из этого, в передающих устройствах, чтобы обеспечить адекватное понимание речи, речевая информация должна передаваться в речевом диапазоне частот.
Структурно-функциональная характеристика
Рецепторный (периферический) отдел слухового анализатора, превращающий энергию звуковых волн в энергию нервного возбуждения, представлен рецепторными волосковыми клетками кортиева органа {орган Корти), находящимися в улитке. Слуховые рецепторы (фонорецепторы) относятся к механорецепторам, являются вторичными и представлены внутренними и наружными волосковыми клетками. У человека приблизительно 3500 внутренних инаружных волосковых клеток, которые расположены на основной мембране внутри среднего канала внутреннего уха.

Внутреннее ухо (звуковоспринимающий аппарат), а также среднее ухо (звукопередающий аппарат) и наружное ухо (звукоулавливающий аппарат) объединяются в понятие орган слуха.
Наружное ухо за счет ушной раковины обеспечивает улавливание звуков, концентрацию их в направлении наружного слухового прохода и усиление интенсивности звуков. Кроме того, структуры наружного уха выполняют защитную функцию, охраняя барабанную перепонку от механических и температурных воздействий внешней среды.
Среднее ухо (звукопроводящий отдел) представлено барабанной полостью, где расположены три слуховые косточки: молоточек, наковальня и стремечко. От наружного слухового прохода среднее ухо отделено барабанной перепонкой. Рукоятка молоточка вплетена в барабанную перепонку, другой его конец сочленен с наковальней, которая, в свою очередь, сочленена со стремечком. Стремечко прилегает к мембране овального окна. Площадь барабанной перепонки (70 мм2) значительно больше площади овального окна (3,2 мм2), благодаря чему происходит усиление давления звуковых волн на мембрану овального окна примерно в 25 раз. Так как рычажный механизм косточек уменьшает амплитуду звуковых волн примерно в 2 раза, то, следовательно, происходит такое же усиление звуковых волн на овальном окне. Таким образом, происходит общее усиление звука средним ухом примерно враз. Если же учитывать усиливающий эффект наружного уха, то эта величина достигает раз. Среднее ухо имеет специальный защитный механизм, представленный двумя мышцами: мышцей, натягивающей барабанную перепонку, и мышцей, фиксирующей стремечко. Степень сокращения этих мышц зависит от силы звуковых колебаний. При сильных звуковых колебаниях мышцы ограничивают амплитуду колебаний барабанной перепонки и движение стремечка, предохраняя тем самым рецепторный аппарат во внутреннем ухе от чрезмерного возбуждения и разрушения. При мгновенных сильных раздражениях (удар в колокол) этот защитный механизм не успевает срабатывать. Сокращение обеих мышц барабанной полости осуществляется по механизму безусловного рефлекса, который замыкается на уровне стволовых отделов мозга. В барабанной полости поддерживается давление, равное атмосферному, что очень важно для адекватного восприятия звуков. Эту функцию выполняет евстахиева труба, которая соединяет полость среднего уха с глоткой. При глотании труба открывается, вентилируя полость среднего уха и уравнивая давление в нем с атмосферным. Если внешнее давление быстро меняется (быстрый подъем на высоту), а глотания не происходит, то разность давлений между атмосферным воздухом и воздухом в барабанной полости приводит к натяжению барабанной перепонки и возникновению неприятных ощущений, снижению восприятия звуков.
Внутреннее ухо представлено улиткой - спирально закрученным костным каналом, имеющим 2,5 завитка, который разделен основной мембраной и мембраной Рейснера на три узких части (лестницы). Верхний канал (вестибулярная лестница) начинается от овального окна и соединяется с нижним каналом (барабанной лестницей) через геликотрему (отверстие в верхушке) и заканчивается круглым окном. Оба канала представляют собой единое целое и заполнены перилимфой, сходной по составу со спинномозговой жидкостью. Между верхним и нижним каналами находится средний (средняя лестница). Он изолирован и заполнен эндолимфой. Внутри среднего канала на основной мембране расположен собственно звуковоспринимающий аппарат - орган Корти (кортиев орган) с рецепторными клетками, представляющий периферический отдел слухового анализатора.
Булава" href="/text/category/bulava/" rel="bookmark">булава , из которой выступает по тончайших ресничек длиной 10 мкм. Обонятельные реснички погружены в жидкую среду, вырабатываемую обонятельными железами. Наличие ресничек в десятки раз увеличивает площадь контакта рецептора с молекулами пахучих веществ.
При спокойном вдохе струя воздуха не попадает в узкую щель между верхней носовой раковиной и носовой перегородкой, где расположена обонятельная область, а потому молекулы пахучих веществ могут проникать в нее лишь с помощью диффузии . Усиленный вдох, а также быстрые, короткие вдыхания, совершаемые при нюхании, вызывают в полости носа вихревые движения воздуха, что способствует проникновению воздуха в обонятельную область. Из полости рта (например, во время еды) молекулы пахучих веществ диффундируют в область носоглотки и легко попадают в полость носа вместе с выдыхаемым воздухом. Чтобы воздействовать на рецепторы, они должны адсорбироваться и раствориться на влажной поверхности обонятельного эпителия.
Чувствительность обонятельных рецепторов необычайно велика. Некоторые вещества, например тринитробутилтолуол, могут быть обнаружены человеком по запаху даже при содержании в литре воздуха миллиардных долей миллиграмма. У многих животных чувствительность обонятельного анализатора во много раз выше, чем у человека.
Наличие огромного количества органических и неорганических пахучих веществ самой различной структуры делает несостоятельными попытки чисто химического объяснения их воздействия на рецепторы. Возможно, что энергия внутримолекулярных колебаний вызывает те физико-химические сдвиги в обонятельном пузырьке, которые приводят к возникновению процесса возбуждения. Такой механизм раздражения, если он действительно существует, был бы сходен с фотохимическим механизмом раздражения светочувствительных элементов сетчатки.
Обонятельные клетки, снабженные рецепторным образованием на конце их периферического отростка, представляют собой первый нейрон проводящих путей обонятельного анализатора. Это типичные биполярные клетки, гомологичные клеткам межпозвоночных узлов спинного мозга. Их аксоны, не покрытые миелиновой оболочкой, образуют до 20 тонких нервных стволиков. Через отверстия решетчатой кости они проходят в полость черепа и проникают в обонятельную луковицу, то есть в передний, утолщенный конец обонятельного тракта. Здесь расположены тела второго нейрона. К дендритам каждого из них подходят конечные ветвления аксонов нескольких биполярных клеток. Аксоны второго нейрона образуют обонятельный тракт и направляются к телам третьего нейрона, расположенном в миндалевидном ядре, в переднем, изогнутом конце аммоновой извилины и в подмозолистой извилине. Аксоны третьего нейрона направляются к корковому отделу обонятельного анализатора.
Помимо указанных основных проводящих путей, достигающих коркового отдела обонятельного анализатора, имеются еще пути, связывающие аксоны второго нейрона с промежуточным мозгом, а также с различными скоплениями серого вещества среднего, заднего и спинного мозга. Через эти пути осуществляются двигательные и сенсорные реакции на раздражение обонятельных рецепторов. По-видимому, уздечка, входящая в состав надбугорья, играет по отношению к рефлексам на раздражение органов обоняния ту же роль, что и четверохолмие по отношению к рефлексам на световые и звуковые раздражения.
Ядро обонятельного анализатора у человека расположено в образованиях старой коры, а именно в глубине борозды аммонова рога. Ядра анализатора обоих полушарий связаны друг с другом проводящими путями. К обонятельному анализатору следует отнести и некоторые соседние образования межуточной коры. Прилегающие участки островковой области, лежащие в глубине сильвиевой борозды, по-видимому, имеют то же значение для обоняния, каким обладают проекционно-ассоциативные поля 18 и 19 для зрительной функции.
Есть основания полагать, что к обонятельному анализатору относится и небольшой участок краевой области, расположенной на внутренней поверхности полушария в виде узкой полоски вдоль мозолистого тела. От коркового отдела обонятельного анализатора идут эфферентные пути к нижележащим отделам мозга, в частности к сосковым телам подбугровой области и к уздечке надбугорья. Через эти пути осуществляются корковые рефлексы на обонятельные раздражения.
Некоторые раздражители, например, ванилин и гваякол, действуют только на обонятельные рецепторы. Многие же другие летучие вещества одновременно раздражают и другие рецепторы.
Так, бензол, нитробензол, хлороформ действуют на вкусовые рецепторы, вследствие чего их запах имеет сладковатый привкус. Хлор, бром, аммиак , формалин возбуждают болевые и тактильные рецепторы слизистой оболочки носа. Ментол, фенол, камфара раздражают холодовые рецепторы, а этиловый спирт - тепловые и болевые. Уксусная кислота действует на вкусовые и болевые рецепторы (отсюда кисловатый и острый запах уксуса) и т. д. Имеются фармакологические и физиологические данные о существовании различных видов рецепторов, обладающих неодинаковой чувствительностью к отдельным запахам. Это свидетельствует о том, что анализ обонятельных раздражений начинается на периферии. Высший анализ и синтез запаховых раздражений происходит в коре больших полушарий.
Сложный характер большинства запаховых ощущений, связанный с одновременным раздражением не только обонятельных, но и других рецепторов, обусловливает тесное взаимодействие корковых отделов трех анализаторов - обонятельного, вкусового, и той части кожного, куда поступают импульсы со слизистой оболочки полости носа. Поэтому в анализе и синтезе запаховых раздражений принимают участие не только упомянутые выше участки коры больших полушарий, но также извилина аммонова рога и нижняя часть постцентральной извилины. Человек не различает отдельные компоненты, входящие в состав сложного запаха. Если смешать два или несколько различно пахнущих веществ, то запах смеси может оказаться либо сходным с запахом одного из них, либо резко отличным от запаха каждой ее составляющей части.
Применяя различные комбинации летучих веществ в строго определенных пропорциях, парфюмеры достигают большого искусства в создании новых запахов. Возможность подавления одного запаха другим используется в целях дезодорации, то есть для нейтрализации запаха зловонных веществ.
Корковые отделы обонятельного анализатора обоих полушарий так тесно связаны между собой, что при чисто обонятельном раздражении человек не различает, в какую половину носовой полости попало летучее вещество. Раздражение других рецепторов полости носа дает локализованные ощущения. Вводя одно запаховое вещество через правую ноздрю, а другое через левую, можно получить подавление одного запаха другим, а также появление совершенно нового запаха. Это показывает, что анализ и синтез обонятельных раздражений в основном происходит не на периферии, а в корковом отделе анализатора. В некоторых случаях удается наблюдать следующее явление -вместо непрерывного ощущения одного и того же запаха смеси возникают чередующиеся ощущения запаха то одного, то другого вещества.
У большинства млекопитающих животных совершенство аналитико-синтетической функции обонятельного анализатора достигает чрезвычайно высоких пределов. У человека в связи с развитием речи и трудовой деятельностью жизненное значение этого анализатора резко снизилось по сравнению со значением зрительного, слухового, тактильного и двигательного анализаторов. Условные рефлексы на действие запаховых раздражителей образуются у человека в неизмеримо меньшем количестве, чем у собаки или кошки; этому соответствует относительно слабое развитие у него коркового отдела обонятельного анализатора. У детей положительные условные рефлексы на запаховые раздражители удается выработать на 5-6-й неделе жизни; образование грубых дифференцировок становится возможным большей частью не ранее начала третьего месяца. Однако тонкие дифференцировки (например, различение разных сортов одеколона) начинают вырабатываться значительно позднее, да и то с большим трудом. Нередко и взрослые люди, несмотря на отсутствие каких-либо нарушений в периферическом отделе анализатора, очень плохо различают запахи.
В тех случаях, когда запаховые раздражения приобретают для человека существенное значение, аналитико-синтетическая деятельность обонятельного анализатора может достигать большого совершенства, вплоть до различения компонентов запаховой смеси. Это наблюдается у некоторых парфюмеров, кулинаров и т. д.
5. Вкусовая рецепция. Виды вкусовых ощущений. Особенности проводникового отдела.
Чувство вкуса связано с раздражением не только химических, но и механических, температурных и даже болевых рецепторов слизистой оболочки полости рта, а также обонятельных рецепторов. Вкусовой анализатор определяет формирование вкусовых ощущений, является рефлексогенной зоной. С помощью вкусового анализатора оцениваются различные качества вкусовых ощущений, сила ощущений, которая зависит не только от силы раздражения, но и от функционального состояния организма.
Структурно-функциональная характеристика вкусового анализатора.
Периферический отдел. Рецепторы вкуса (вкусовые клетки с микроворсинками) - это вторичные рецепторы, они являются элементом вкусовых почек, в состав которых входят также опорные и базальные клетки. Во вкусовых почках обнаружены клетки, содержащие серотонин, и клетки, образующие гистамин. Эти и другие вещества играют определенную роль в формировании чувства вкуса. Отдельные вкусовые почки являются полимодальными образованиями, так как могут воспринимать различные виды вкусовых раздражителей. Вкусовые почки в виде отдельных включений находятся на задней стенке глотки, мягком нёбе, миндалинах, гортани, надгортаннике и входят также в состав вкусовых сосочков языка как органа вкуса.
Периферический отдел вкусового анализатора представлен вкусовыми луковицами, которые расположены главным образом в сосочках языка. Вкусовые клетки усеяны на своем конце микроворсинками, которые называют еще вкусовыми волосками. Они выходят на поверхность языка через вкусовые поры.
На вкусовой клетке имеется большое число синапсов, которые образуют волокна барабанной струны и языкоглоточного нерва. Волокна барабанной струны (ветвь язычного нерва) подходят ко всем грибовидным сосочкам, а волокна языкоглоточного нерва - к желобоватым и листовидным. Корковый конец вкусового анализатора находится в гиппокампе , парагиппокамповой извилине и в нижней части заднецентральной извилины.
Вкусовые клетки непрерывно делятся и непрерывно гибнут. Особенно быстро происходит замещение клеток, расположенных в передней части языка, где они лежат более поверхностно. Замена клеток вкусовой почки сопровождается образованием новых синаптических структур
Проводниковый отдел. Внутрь вкусовой почки входят нервные волокна, которые образуют рецепторно-афферентные синапсы. Вкусовые почки различных областей полости рта получают нервные волокна от разных нервов: вкусовые почки передних двух третей языка - от барабанной струны, входящей в состав лицевого нерва; почки задней трети языка, а также мягкого и твердого нёба, миндалин - от языкоглсточного нерва; вкусовые почки, расположенные в области глотки, надгортанника и гортани, - от верх-пегортанного нерва, являющегося частью блуждающего нерва.
Эти нервные волокна являются периферическими отростками биполярных нейронов, расположенных в соответствующих чувствительных ганглиях, представляющих первый нейрон проводникового отдела вкусового анализатора. Центральные отростки этих клеток входят в состав одиночного пучка продолговатого мозга, ядра которого представляют второй нейрон. Отсюда нервные волокна в составе медиальной петли подходят к зрительному бугру (третий нейрон).
Центральный отдел. Отростки нейронов таламуса идут в кору больших полушарий (четвертый нейрон). Центральный, или корковый, отдел вкусового анализатора локализуется в нижней части соматосенсорной зоны коры в области представительства языка. Большая часть нейронов этой области мультимодальна, т. е. реагирует не только на вкусовые, но и на температурные, механические и ноцицептивные раздражители. Для вкусовой сенсорной системы характерно то, что каждая вкусовая почка имеет не только афферентные, но и эфферентные нервные волокна, которые подходят к вкусовым клеткам из ЦНС, благодаря чему обеспечивается включение вкусового анализатора в целостную деятельность организма.
Механизм вкусового восприятия. Чтобы возникло вкусовое ощущение, раздражающее вещество должно находиться в растворенном состоянии. Сладкое или горькое вкусовое вещество, растворяющееся в слюне до молекул, проникает в поры вкусовых луковиц, вступает во взаимодействие с гликокаликсом и адсорбируется на клеточной мембране микроворсинки, в которую встроены «сладкочувствующие» или «горькочувствующие» рецепторные белки. При воздействии соленых или кислых вкусовых веществ изменяется концентрация электролитов около вкусовой клетки. Во всех случаях повышается проницаемость клеточной мембраны микроворсинок, возникает движение ионов натрия внутрь клетки, происходят деполяризация мембраны и образование рецепторного потенциала, который распространяется и по мембране, и по микротубулярной системе вкусовой клетки к ее основанию. В это время во вкусовой клетке образуется медиатор (ацетилхолин, серотонин, а также, возможно, гормоноподобные вещества белковой природы), который в рецепторно-афферентном синапсе ведет к возникновению генераторного потенциала, а затем потенциала действия во внесинаптических отделах афферентного нервного волокна.
Восприятие вкусовых раздражителей. Микроворсинки вкусовых клеток являются образованиями, непосредственно воспринимающими вкусовой раздражитель. Мембранный потенциал вкусовых клеток колеблется от -30 до -50 мВ. При действии вкусовых раздражителей возникает рецепторный потенциал величиной от 15 до 40 мВ. Он представляет собой деполяризацию поверхности вкусовой клетки, которая является причиной возникновения в волокнах барабанной струны и языкоглоточного нерва генераторного потенциала, переходящего по достижении критического уровня в распространяющиеся импульсы. С рецепторной клетки возбуждение передается через синапс на нервное волокно с помощью ацетилхолина. Некоторые вещества, как например СаС12, хинин, соли тяжелых металлов, вызывают не первичную деполяризацию, а первичную гиперпрляризацию. Ее возникновение связано с осуществлением отрицательных отвергаемых реакций . Распространяющихся импульсов при этом не возникает.
В отличие от обонятельных вкусовые ощущения легко могут быть объединены в группы по сходным признакам. Различают четыре основных вкусовых ощущения - сладкое, горькое, кислое и соленое, которые в своих сочетаниях могут давать многообразные оттенки вкусовых качеств.
Ощущение сладкого вызывается содержащимися в пищевых веществах углеводами (двухатомные и многоатомные спирты, моносахариды и т. д.); ощущение горького - воздействием на вкусовые рецепторы различных алкалоидов ; ощущение кислого возникает от действия растворенных в воде различных кислот; ощущение соленого вызывается поваренной солью (хлористый натрий) и другими соединениями хлора.
6. Кожный анализатор: виды рецепции, проводниковый отдел, представительство в коре больших полушарий.
К кожному анализатору относят совокупность анатомических образований, согласованной деятельностью которых определяются такие виды кожной чувствительности, как чувство давления, растяжения, прикосновения, вибрации, тепла, холода и боли. Все рецепторные образования кожи в зависимости от их структуры делятся на две группы: свободные и несвободные. Несвободные в свою очередь подразделяются на инкапсулированные и неинкапсулированные.
Свободные нервные окончания представлены конечными разветвлениями дендритов сенсорных нейронов. Они теряют миелин, проникают между клетками эпителия и располагаются в эпидермисе и дерме. В некоторых случаях конечные разветвления осевого цилиндра окутывают измененные эпителиальные клетки, образуя осязательные мениски.
Несвободные нервные окончания состоят не только из ветвлений волокна, потерявшего миелин, но и из клеток глии. К несвободным инкапсулированным рецепторным образованиям кожи относятся пластичные тельца, или тельца Фатера-Пачини, заметные невооруженным глазом (например, на разрезе кожи пальцев руки), в жировой клетчатке. Прикосновение воспринимается осязательными тельцами (тельца Мейснера, колбы Краузе и др.) сосочкового слоя собственно кожи, осязательными дисками росткового слоя эпидермиса. Корни волос оплетены нервными манжетками.
Густота расположения рецепторов в коже разных участков тела неодинакова и функционально обусловлена. Заложенные в коже рецепторы служат периферическими частями кожного анализатора, имеющего благодаря её протяженности существенное значение для организма.
Возбуждение от рецепторов кожного анализатора направляется в центральную нервную систему по тонкому и клиновидному пучкам. Кроме того, импульсы от рецепторов кожи проходят по спинно-бугорному пути и троичной петле, а от проприорецепторов – по спиномозжечковым путям.
Тонкий пучок несёт импульсы от тела ниже 5 грудного сегмента, а клиновидный пучок – от верхней части туловища и рук. Эти пути образованы нейритами чувствительный нейронов, тела которых лежат в спинномозговых узлах, а дендриты оканчиваются рецепторами кожи. Пройдя весь спинной мозг и заднюю часть продолговатого, волокна тонкого и клиновидного пучков оканчиваются на нейронах тонного и клиновидного ядер. Волокна тонкого и клиновидного ядер идут по двум направлениям. Одни – под названием наружных дугообразных волокон – переходят на противоположную сторону, где в составе нижних ножек мозжечка оканчиваются на клетках коры его червя. Нейриты последних связывают кору червя с ядрами мозжечка. Волокна клеток этих ядер в составе нижних ножек мозжечка направляются к преддверным ядрам моста.
Другая, большая часть волокон клеток тонкого и клиновидного ядер спереди от центрального канала продолговатого мозга совершает перекрест и образует медиальную петлю. Последняя идёт через продолговатый мозг, покрышки моста и среднего мозга и заканчивается в вентральном ядре зрительного бугра. Волокна нейронов зрительного бугра идут в составе таламической лучистости к коре центральных областей большого полушарий.
Спинно-бугорный путь проводит возбуждение от рецепторов, раздражение которых вызывает болевые и температурные ощущения. Тела чувствительных нейронов этого пути залегают в спинномозговых ганглиях. Центральные волокна нейронов входят в составе задних корешков в спинной мозг, где оканчиваются на телах вставочных нейронов задних рогов. Отростки клеток задних рогов переходят на противоположную сторону и в глубине бокового канатика соединяются в спинно-бугорный путь. Последний проходит спинной мозг, покрышки продолговатого мозга, моста и ножек мозга и оканчивается на клетках вентрального ядра зрительного бугра. Волокна этих нейронов идут в составе таламической лучистости к коре, где и оканчиваются, главным образом в заднецентральной области.
Троичная петля передаёт импульсы от рецепторов кожи головы. Чувствительными нейронами служат клетки троичного узла. Периферические волокна этих клеток проходят в составе трёх ветвей троичного нерва, иннервирующих кожу лица. Центральные волокна чувствительных нейронов выходят из узла в составе чувствительного корешка троичного нерва и проникают в мозг в том месте, где он переходит в средние ножки мозжечка. В мосту эти волокна делятся Т-образно на восходящие и нисходящие ветви (спинномозговой путь), которые оканчиваются на нейронах, образующих в покрышке моста основное сенсорное ядро троичного нерва, а в продолговатом и спинном мозге – ядро его спинномозгового пути. Центральные волокна этих ядер совершают перекрест в верхней части моста и в качестве троичной петли проходят по покрышке среднего мозга до зрительного бугра, где оканчиваются самостоятельно или вместе с волокнами медиальной петли на клетках его вентрального ядра. Отростки нейронов этого ядра направляются в составе таламической лучистости к коре нижней части заднецентральной области, где главным образом и локализуется кожный анализатор головы.
7. Орган равновесия: значение в жизни человека и животных. Особенности рецепторного аппарата.
Вестибулярный анализатор или орган равновесия обеспечивает ощущение положения и перемещения человеческого тела или его частей в пространстве, а также обусловливает ориентацию и поддержание позы при всех возможных видах деятельности человека

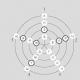
Рис 17 Строение и расположение лабиринта и рецепторов отолитового аппарата:
1, 2, 3 - соответственно горизонтальный, фронтальный и сагиттальный полукругом каналы; 4,5 - отолиты аппарат: овальный (4) и круглый (5) мешочки; 6,7 - нервные ганглии, 8 - вестибуло-кохлеарный нерв (Ш ра черепно-мозговых нервов), 9 - отолиты; 10-желеобразная масса, 11 - волоски, 12 - рецепторные волосковые клетки, 13 - опорные клетки, 14 - нервные волокна
Периферический (рецепторный) отдел вестибулярного анализатора расположен, как и внутреннее ухо, в лабиринтах пирамиды височной кости Лежит он в так называемом вестибулярном а аппарате (рис 17) и состоит из преддверия (отолитового органа) и трех полукружных каналов, расположенных втроем взаимно перпендикулярных плоскостях: горизонтальной, фронтальной (слева на право), и с агитальний (переднее-задней) преддверие или переддверя состоит, как указывалось, из двух перепончатых мешочков: круглого, расположенного ближе к завитка внутреннего уха и овального (пестика), располо шованого ближе к полукружных каналов перепончатого части полукружных каналов соединены пятью отверстиями с пестиком преддверия Начальный конец каждого полукружных каналов имеет расширение, что называется ампулой Все перепончатые части вестибулярного анализатора заполнены эндолимфой Вокруг перепончатого лабиринта, (между ним и его костным футляром) находится перилимфой, которая переходит также в пе рилимфу внутреннего уха На внутренней поверхности мешочков имеются небольшие возвышения (пятна) где именно и расположены рецепторы равновесия, или отолитовой аппарат, который размещен полувертикальном в овальном Мише чку и горизонтально в круглом мешочке В отолитового аппарата находятся рецепторные волосковые клетки (механорецепторы), имеющие на своей вершине волоски (реснички) двух типов, много тонких и коротких стереоцилиив и один более толстый и длинный волосок, произрастающего на периферии и называется киноцилий Рецепторные волосковые клетки пятен на поверхности мешочков преддверия собраны в группы, называемые мака ли Киноцилии всех волосковых клеток погружены в студенистую массу расположенной над ними так называемой отолитовой мембраны, содержащей многочисленные кристаллы фосфата и карбоната кальция, называемые отолиты (в дословном переводе - ушные камни) Концы стереоцилиив волосковых клеток макулы свободно подпирают и удерживают на себе отолитовой мембрану (рис. 18).
Благодаря отолиты (твердым включением), плотность отолитовой мембраны выше плотность среды, что его окружает Под действием силы тяжести гравитации или ускорения, отолитовой мембрана смещается относительно рец цепторних клеток, волоски (киноцилии) этих клеток сгибаются и в них возникает возбуждение Таким образом, отолитового аппарата каждое мгновение контролирует расположение тела относительно силы тяжести, определяет, в котором пол оженни в пространстве (в горизонтальном или в вертикальном) находится тело, а также реагирует на прямолинейные ускорения при вертикальных и горизонтальных движениях тела Порог чувствительности отолитового аппарата до прямолинейных ускорений равна 2-20 см / сек, а порог распознавания наклона головы в сторону составляет 1 °; вперед и назад - около 2 ° При сопутствующих раздражениях (при воздействии вибрации, колебания, тряски) чувствительность вестибулярного анализатора снижается (например, вибрации транспорта могут повышать порог распознавания наклона головы вперед и назад до 5 °, а в сторону-до 10 ° 10°).
Второй частью вестибулярного аппарата есть три полукружных канала, каждый диаметром около 2 мм На внутренней поверхности ампул полукружных каналов (рис 18) расположены гребешки, на вершине которых волосковые клетки сгруппированы в кристы, над которыми расположена студенистая масса с отолитов, что здесь называется листовидной мембраной или купулою Киноцилии волосковых клеток крист, так как это было описано и для о толитового аппарата мешочков преддверия, погружены в купулу и возбуждаются от движений эндолимфы, возникающие при перемещениях тела в пространстве При этом наблюдается движение волосков - стереоцилиив в сторону киноцилиив Возникает рецепторный потенциал действия волосковых клеток, выделяется медиатор ацетилхолин, который и стимулирует синаптические окончания вестибулярного нерва Если смещение стереоцилиив направлено в пр отилежний от киноцилий сторону, то активность вестибулярного нерва наоборот снижается Дня волосковых клеток полукружных каналов адекватным раздражителем является ускорение или замедление вращения в определенных пло щинах Дело в том, что эндолимфа полукружных каналов имеет такую??же плотность, как и купула ампул и поэтому прямолинейные ускорения не влияют на положение волосков волосковых клеток и купулы При оборота УНИ головы или тела возникают угловые ускорения и тогда купула начинает двигаться, возбуждая рецепторные клетки Порог распознавания вращения для рецепторов полукружных каналов составляет примерно 2-3 ° / это 2-3 °/сек.
К рецепторов вестибулярного аппарата подходят периферийные волокна биполярных нейронов вестибулярного ганглия, ию расположенный во внутреннем ухе (первые нейроны) Аксоны этих нейронов сплетаются вместе м с нервными волокнами от рецепторов внутреннего уха и образуют единый вестибуло-кохлеарний или присинковых-улитковый нерв (VIII пара черепно-мозговых нервов) Импульсы возбуждения о положении те а в пространстве этим нервом поступают в продолговатого мозга (второй нейрон), в частности в вестибулярный центр, куда также приходят нервные импульсы от рецепторов мышц и суставов Третий нейрон расположен и в ядрах зрительных бугорков среднего мозга, которые в свою очередь соединены нервными путями с мозжечком (отделом мозга, обеспечивающей координацию движений), а также с подкорковыми образованиями и корой головного ого мозга (центрами движения, письма, речи, глотания и т. д.) Центральный отдел вестибулярного анализатора локализуется в височной доле головного мозгаозку.
При возбуждении вестибулярного анализатора возникают соматические реакции (на основе вестибуло-спинальных нервных связей), способствующих перераспределению тонуса мышц и постоянной поддержке равновесия тела в пространстве Рефлексы, обеспечивающие равновесие тела делятся на статические (вне стояния, сидения и др.) и статокинетического Примером статокинетического рефлекса может быть вестибулярный нистагм глаз ность АГМ возникает в условиях быстрого перемещения тела или его вращения и заключается в том, что глаза сначала медленно движутся в сторону, противоположную направлению перемещения или вращения, а затем быстрым движением в обратной ротному направлении перескакивают на новую точку фиксации зрения Реакции такого типа обеспечивают возможность обозрения пространства в условиях движения тел тіла.
Благодаря связям вестибулярных ядер с мозжечком обеспечиваются все подвижные реакции и реакции по координации движений, в том числе при выполнении трудовых операций или спортивных упражнений Поддержанию рав веса также способствуют зрение и мышечно-суставная рецепцийя.
Связь вестибулярных ядер с вегетативной нервной системой обусловливает вестибуло-вегетативные реакции сердечно-сосудистой системы, желудочно-кишечного тракта и других органов Такие реакции могут проявляться я в изменениях сердечного ритма, тонуса сосудов, артериального давления, могут возникать тошнота и рвота (например, как это бывает при длительной и сильной действия специфических раздражителей движения транспорта на вестибюль овражный аппарат, что приводит к укачивания).
Формирование вестибулярного аппарата у детей заканчивается раньше других анализаторов У новорожденного ребенка этот орган функционирует почти так же, как и у взрослого человека Тренировка двигательных качеств у ди итей с самого раннего детства способствует оптимизации развития вестибулярного анализатора и, как результат, разнообразит их двигательные возможности, до феноменальных (например, упражнения цирковых акробатов гимнастов и др. н.).
При действии стимула на рецепторную мембрану происходит повышение проницаемости для ионов натрия или кальция, ионы поступают внутрь нервного окончания, мембрана деполяризуется и формируется рецепторный потенциал.
Рецепторный потенциал обладает всеми свойствами местного возбуждения (зависит от силы раздражителя, способен к суммации, распространяется с затуханием). Затем рецепторный потенциал с помощью местных токов вызывает генерацию ПД в афферентном волокне. Частота ПД зависит от амплитуды рецепторного потенциала.
Открываются Ca/Na каналы → поступают внутрь клетки по пассивному градиенту концентрации → деполяризация мембраны.
Формирование локальных токов выходящего направления → деполяризация до КУД → генерация ПД.
Механизмы возбуждения вторичночувствующих рецепторов
В рецепторной клетке под действием раздражителя открываются натриевые или кальциевые каналы, что приводит к возникновению рецепторного потенциала. Возбуждение клетки вызывает секрецию медиатора (ацетилхолин, глутаминовая кислота), и на мембране чувствительного нейрона формируется генераторный потенциал.
Генераторный потенциал с помощью местных токов действует на мембрану афферентного волокна, где и возникает ПД
Рецепторныйпотенциал возникает при раздражении рецептора как результат деполяризации и повышения проводимости участка его мембраны, который называется рецептивным. Возникший в рецептивных участках мембраны рецепторный потенциал электротонически распространяется на аксонный холмик рецепторного нейрона, где возникает генераторный потенциал . Возникновение генераторного потенциала в области аксонного холмика объясняется тем, что этот участок нейрона имеет более низкие пороги возбуждения и потенциал действия в нем развивается раньше, чем в других частях мембраны нейрона. Чем выше генераторный потенциал, тем интенсивнее частота разрядов распространяющегося потенциала действия от аксона к другим отделам нервной системы. Следовательно, частота разрядов рецепторного нейрона зависит от амплитуды генераторного потенциала.
Афферентное звено рефлекторной дуги представлено сенсорными нейронами, тела которых находятся в чувствительных ганглиях спинномозговых нервов или соответствующих черепномозговых нервах.
Центральное звено рефлекса представлено интернейронами, которые образуют малые возбуждающие и тормозные нейронные сети.
Эфферентное звено представлено одним (для соматических рефлексов) или двумя (для вегетативных рефлексов) нейронами.
Эффекторный орган в соматических рефлексах - это скелетные мышцы, в вегетативных - гладкие мышцы, железы, кардиомиоциты.
Морфофункциональная характеристика симпатического отдела вегетативной нервной
Системы.
Центры СНС локализованы в торако-люмбальном отделе спинного мозга.
Преганглионарные нейроны лежат в боковых рогах от последнего шейного до 4-го поясничного сегмента спинного мозга (С8, Th1-Th12, L1-L4).
Имеется 2 типа ганглиев в СНС:
1. паравертебральные ганглии являются парными и образуют симпатическую нервную цепочку по обе стороны от спинного мозга (симпатический ствол)
2. превертебральные ганглии - непарные, их три (солнечное сплетение, верхний и нижний брыжеечные узлы).
Медиаторы и реактивные системы
ñ Преганглионарные нейроны - холинергические (Ах).
ñ Постганглионарные нейроны - адренергические (Над),
ñ Передача возбуждения на орган осуществляется с помощью альфа- и бета-адренорецепторов, которые являются метаботропными рецепторами.
При возбуждении альфа-адренорецепторов активируется мембранный фермент фосфолипаза С и образуется два вторичных посредника: инозитолтрифосфат (ИТФ) и диацилглицерол (ДАГ) .
При возбуждении бета-адренорецепторов активируется мембранный фермент аденилатциклаза, что приводит к образованию вторичного посредника ц-АМФ . Следствием активации адренорецепторов может быть изменение как натриевой, так и калиевой проводимости мембраны эффекторных клеток.
При действии стимула на рецептор происходит изменение пространственной структуры белковых молекул, встроенных в мембрану рецепторной клетки. Это приводит к изменению проницаемости мембраны главным образом для ионов натрия и в некоторой степени калия. В результате возникает ионный ток, который изменяет потенциал покоя и вызывает генерацию рецепторного потенциала, который является первичным электрическим ответом рецептора на действие стимула.
В первично чувствующих рецепторах рецепторный потенциал достигает критического уровня и приводит к генерации распространяющихся потенциалов действия, которые без затухания передаются по афферентным нервным волокнам в вышележащие уровни центральной нервной системы.
Во вторично чувствующих рецепторах рецепторный потенциал вызывает выделение медиатора (ацетилхолина) из пресинаптического окончания рецепторной клетки. Медиатор воздействует на постсинаптическую мембрану чувствительного нейрона и вызывает ее деполяризацию, т.е. развивается постсинаптический потенциал. Постсинаптический потенциал первого чувствительного нейрона называется генераторным потенциалом. В отличие от потенциалов действия, генерация которых подчиняется закону «все или ничего», величина генераторного потенциала зависит от амплитуды рецепторного потенциала и пропорциональна величине стимула. Когда генераторный потенциал достигает определенного критического уровня, он формирует на чувствительном нервном окончании серию распространяющихся нервных импульсов. Таким образом, в первично чувствующих рецепторах рецепторный потенциал является в тоже время и генераторным потенциалом, а во вторично чувствующих рецепторный потенциал и генераторный потенциал разобщены.
Характеристика рецепторов
Рецепторы обладают способностью не только воспринимать, но и усиливать сигнал за счет собственной внутренней энергии – энергии метаболических процессов. Следовательно, количество энергии, затрачиваемое рецепторами на восприятие и кодирование его в серии нервных импульсов, может значительно отличаться от энергии стимула.
Деятельность разнообразных рецепторных аппаратов направлена, в конце концов, на обеспечение эффективной работы сенсорных систем. Это предполагает, несмотря на вариативность в строении и механизмах работы конкретных рецепторов, выделить особенности, характерные для всех видов.
1. Ритмическая активность . Многим рецепторам свойственна ритмическая активность в состоянии покоя – спонтанная или фоновая импульсация. Фоновая активность позволяет рецепторам оценивать не только возбуждающие, но и тормозные воздействия, т. е. передавать сведения о сигнале не только в виде учащения, но и в виде урежения потока импульсов. Кроме того, фоновая импульсация позволяет поддерживать высокую возбудимость рецепторов.
2. Адаптация рецепторов . Это процесс уменьшения активности рецепторов по мере действия раздражителя с постоянными физическими характеристиками. Наиболее быстро адаптирующиеся рецепторы – механорецепторы кожи. Практически отсутствует способность к адаптации у проприорецепторов.
При длительном неизменном раздражении адаптация проявляется в снижении, а затем в полном исчезновении рецепторного потенциала. По своему характеру адаптация может быть полной или неполной, а также быстрой и медленной. Как правило, быстрая адаптация бывает полной, а медленная – неполной. Исчезновение рецепторного потенциала при адаптации рецептора к действию постоянного стимула приводит к потере определённой доли информации. Но рецептор сохраняет способность мгновенно реагировать на любое изменение параметров раздражения. Благодаря способности рецепторов к адаптации значительная доля информации не достигает сознания и таким образом не перегружает высшие отделы центральной нервной системы избыточной информацией.
3. Направленная чувствительность. У многих рецепторов максимальное возбуждение отмечается при определенном направлении раздражающего возбуждения, а при другом направлении рецептор возбуждается минимально или тормозится.
4. Эфферентная регуляция. Рецепторы находятся под тоническим тормозным влиянием со стороны вышележащих нервных структур. Это торможение стабилизирует мембранный потенциал около определенной величины, благодаря чему генераторный потенциал не достигает критического уровня, и потенциалы действия в рецепторе не возникают. Роль направленной чувствительности состоит в формировании на высших этажах сенсорной системы специализированных нейронов-детекторов направленного движения.
Вопросы и задания для самоконтроля
1. Дайте определение понятия «рецептор», назовите функции рецепторов.
2. Рассмотрите различные классификации рецепторов.
3. Каков механизм возбуждения первично и вторично чувствующих рецепторов? Приведите примеры первично и вторично чувствующих рецепторов.
4. Чем отличаются друг от друга быстро и медленно адaптирующиеся рецепторы?
5. К какому виду рецепторов в рамках различных классификаций относятся зрительные, слуховые, обонятельные, вкусовые рецепторы, рецепторы мышц и другие?
6. Какие процессы лежат в основе возникновения рецепторного потенциaла?
7. В чем разница между рецепторным потенциалом, генераторным потенциaлом и потенциaлом действия?
8. Назовите характерные особенности рецепторных элементов сенсорных систем.
В случае первичных рецепторов, действие раздражителя воспринимается окончанием чувствительного нейрона. Действующий раздражитель может вызывать гиперполяризацию или деполяризацию поверхностной мембраны рецепторы в основном за счет изменения натриевой проницаемости. Повышение проницаемости к ионам натрия приводит к деполяризации мембраны и на мембране рецептора возникает рецепторный потенциал. Он существует до тех пор, пока действует раздражитель.
Рецепторный потенциал не подчиняется закону «Все или ничего», его амплитуда зависит от силы раздражителя. У него нет периода рефрактерности. Это позволяет суммироваться рецепторным потенциалам при действии последующих раздражителей. Он распространяется мелено, с угасанием. Когда рецепторный потенциал достигает критической пороговой величины, он вызывает появление потенциала действия в ближайшем перехвате Ранвье. В перехвате Ранвье возникает потенциал действия, который подчиняется закону «Все или ничего» Этот потенциал будет распространяющимся.
Во вторичном рецепторе действие раздражителя воспринимается рецепторной клеткой. В этой клетке возникает рецепторный потенциал, следствием которого будет являться выделение медиатора из клетки в синапс, который действует на постсинаптическую мембрану чувствительного волокна и взаимодействие медиатора с рецепторами приводит к образованию другого, локального потенциала, который называют генераторным . Он по своим свойства идентичен рецепторным. Его амплитуда определяется количеством выделившегося медиатора. Медиаторы – ацетилхолин, глутамат.
Потенциалы действия возникают периодически, т.к. для них характерен период рефрактерности, когда мембрана утрачивает свойство возбудимости. Потенциалы действия возникают дискретно и рецептор в сенсорной системе работает, как аналогово-дискретный преобразователь. В рецепторах наблюдается приспособление – адаптация к действию раздражителей. Есть быстроадаптирующиеся, есть медленно адаптирующиеся. При адаптация снижается амплитуда рецепторного потенциала и число нервных импульсов, которые идут по чувствительному волокну. Рецепторы кодируют информацию. Оно возможно по частоте потенциалов, по группировки импульсов в отдельные залпы и интервалами между залпами. Кодирование возможно по числу активированных рецепторов в рецептивном поле.
Порог раздражения и порог развлечения.
Порог раздражения – минимальная сила раздражителя, которая вызывает ощущение.
Порог развлечении – минимальная сила изменения раздражителя, при которой возникает новое ощущение.
Волосковые клетки возбуждаются при смещении волосков на 10 в -11 метра – 0,1 амстрема.
В 1934 году Вебер сформулировал закон, устанавливающий зависимость между первоначальной силой раздражения и интенсивностью ощущения. Он показал, что изменение силы раздражителя, етсь величина постоянная
∆I / Io = К Io=50 ∆I=52,11 Io=100 ∆I=104,2
Фехнер определили, что ощущение прямопропорционально логарифму раздражения
S=a*logR+b S-ощущение R- раздражение
S=KI в Aстепени I – сила раздражения, К и А – константы
Для тактильных рецепторов S=9,4*I d 0,52
В сенсорных системах есть рецепторы саморегуляции чувствительности рецепторов.
Влияние симпатической системы – симпатическая система повышает чувствительность рецепторов к действию раздражителей. Это полезно в ситуации опасности. Повышает возбудимость рецепторов – ретикулярная формация. В составе чувствительных нервов обнаружены эфферентные волокна, которые могут изменять чувствительность рецепторов. Такие нервные волокна есть в слуховом органе.
Сенсорная система слуха
У большинства людей, живущих в современной остановке слух прогрессивно падает. Это происходит с возрастом. Этому способствует загрязнение звуками окружающей среды – автотранспорт, дискотека и др. Изменения в слуховом аппарате становятся не обратимыми. Уши человека содержат 2 чувствительных органа. Слух и равновесие. Звуковые волны распространяются в форме сжатий и разряжений в упругих средах и при этом распространение звуков в плотных средах идет лучше, чем в газах. Звук обладает 3мя важными свойствами – высотой или частотой, мощностью, или интенсивностью и тембром. Высота звука зависит от частоты колебаний и ухо человека воспринимает с частотой от 16 до 20000 Гц. С максимальной чувствительностью от 1000 о 4000 Гц.
Основная частота звука гортани мужчины – 100 Гц. Женщины - 150 Гц. При разговоре возникают дополнительные высокочастотные звуки в форме шипения, свиста, которые исчезают при разговоре по телефону и это делает речь понятнее.
Мощность звука определяется амплитудой колебаний. Мощность звука выражают в Дб. Мощность представляет собой логарифмическую зависимость. Шепотная речь – 30 Дб, нормальная речь – 60-70 Дб. Звук транспорта – 80, шум мотора самолета – 160. Мощность звука 120 Дб вызывает дискомфорт, а 140 приводят к болезненным ощущениям.
Тембр определяется вторичными колебаниями на звуковых волнах. Упорядоченные колебания – создают музыкальные звуки. А беспорядочные колебания вызывают просто шум. Одна и та же нот звучит по разному на разных инструментах из за разных дополнительных колебаний.
Ухо человека имеет 3 составные части – наружное, среднее и внутренне ухо. Наружное ухо представлено ушной раковиной, которое действует как звука улавливающая воронка. Ухо человека менее совершенно улавливает звуки, чем у кролика, лошади, которые умеют управлять своими ушами. В основе ушной раковины – хрящ, за исключением мочки уха. Хрящевая ткань придает эластичность и форму уху. Если хрящ повреждается, то он восстанавливается разрастаясь. Наружный слуховой проход S образной формы – внутрь, вперед и вниз, длина 2,5 см. Слуховой проход покрыт кожей с малой чувствительностью наружной части и высокой чувствительностью внутренней. В наружной части слухового прохода имеются волосы, которые предупреждают попадание в слуховой проход частиц. Железы слухового прохода вырабатывают желтую смазку, которая тоже предохраняет слуховой проход. В конце прохода – барабанная перепонка, которая состоит из фиброзных волокон, покрытых снаружи кожей, а внутри – слизистой. Барабанная перепонка отделяет среднее от наружного уха. Она колеблется с частотой воспринимаемого звука.
Среднее ухо представлено барабанной полостью, объем которой равен примерно 5-6 капель воды и барабанная полость заполнена водухом, выстлана слизистой оболочкой и содержит 3 слуховые косточки: молоточек, наковальня и стремечко.среднее ухо сообщается с носоглоткой с помощью евстахиевой трубы. В состоянии покоя просвет евстахиевой трубы закрыт, что выравнивает давление. Воспалительные процессы, приводящие к воспалению этой трубы вызывают ощущение заложенности. Среднее ухо отделено от внутреннего овальным и круглым отверстием. Колебания барабанной перепонки через систему рычагов передаются стремечком на овальное окно, причем наружное ухо осуществляет передачу звуков воздушным способом.
Имеется различие площади барабанной перепонки и овального окна(площадь барабанной перепонки равна 70мм в кв. а у овального окна- 3.2мм в кв). При передаче колебания с перепонки на овальное окно амплитуда уменьшается а сила колебаний увеличивается в 20-22 раза. В частотах до 3000 Гц передается 60% Е на внутреннее ухо. В среднем ухе имеется 2 мышцы изменяющие колебания: мышца напрягающая барабанную перепонку(прикрепляется к центральной части барабанной перепонки и к рукоятке молоточка)- при увеличении силы сокращения уменьшается амплитуда; мышца стремечка- ее сокращения ограничивают колебания стремечка. Эти мышцы предупреждают травмы барабанной перепонки. Кроме воздушной передачи звуков есть и костная передача, но это сила звука не в состоянии вызвать колебания костей черепа.
Внутрее ухо
внутреннее ухо представляет собой лабиринт, состоящий из взаимосвязанных трубочек и расширений. Во внутреннем ухе располагается орган равновесия. Лабиринт имеет костную основу, а внутри располагается перепончатый лабиринт и там находится эндолимфа. К слуховой части относится улитка, она образует 2.5 оборота вокруг центральной оси и делится на 3 лестницы: вестибулярная, барабанная и перепончатая. Вестибулярный канал начинается мембраной овального окна, а заканчивается круглым окном. На вершине улитки эти 2 канала сообщаются с помощью геликокрема. А оба этих канала заполнены перилимфой. В среднем перепончатом канале располагается звуковоспринимающий аппарат - кортиев орган. Основная мембрана построена из эластических волокон, которые начинаются у основания(0.04мм) и до вершины (0.5мм). К вершине плотность волокон уменьшается в 500 раз. На основной мембране располагается кортиев орган. Он построен из 20-25 тысяч специальных волосковых клеток, расположенных на поддерживающих клетках. Волосковые клетки лежат в 3-4 ряда(наружный ряд) и в один ряд(внутренний). На вершине волосковых клеток имеются стереоцили или киноцили- самые большие стереоцили. К волосковым клеткам подходят чувствительные волокна 8 пары ЧМН от спирального ганглия. При этом 90% выделенных чувствительных волокон оказываются на внутренних волосковых клетках. На одну внутреннюю волосковую клетку конвергирует до 10 волокон. А в составе нервных волокон есть и эфферентные(оливо-улиточный пучок). Они образуют тормозные синапсы на чувствительных волокнах от спирального ганглия и иннервирует наружные волосковые клетки. Раздражение кортиевого органа связано с передачей колебаний косточек на овальное окно. Низкочастотные колебания распространяются от овального окна до вершины улитки (вовлекается вся основная мембрана).при низких частотах наблюдается возбуждение волосковых клеток лежащих на вершине улитки. Изучением распространения волн в улитке занимался Бекаши. Он обнаружил, что с увеличением частоты вовлекается меньший по протяженности столб жидкости. Высокочастотные звуки не могут вовлечь весь столб жидкости, поэтому чем больше частота, тем меньше колеблется перилимфа. Колебания основной мембраны могут возникать при передаче звуков через перепончатый канал. При колебании основной мембраны происходит смещение волосковых клеток вверх, что вызывает деполяризацию, а если вниз- волоски отклоняются внутрь, что приводит к гиперполяризации клеток. При деполяризации волосковых клеток открываются Са-каналы и Са способствует потенциалу действия, который несет информацию о звуке. Наружные слуховые клетки имеют эфферентную иннервацию и передача возбуждения идет с помощью Асh на наружных волосковых клетках. Эти клетки могут изменять свою длину: они укорачиваются при гиперполяризации и удлиняются при поляризации. Изменение длины наружных волосковых клеток влияет на колебательный процесс, что улучшает восприятие звука внутренними волосковыми клетками. Изменение потенциала волосковых клеток связано с ионным составом эндо- и перилимфы. Перилимфа напоминает ликвор, а эндолимфа имеет высокую концентрацию К(150 ммоль). Поэтому эндолимфа приобретает положительный заряд к перилифме.(+80мВ). Волосковые клетки содержат много К; они имеют мембранный потенциал и отрицательно заряженный внутри и положительный снаружи(МП=-70мВ), а разница потенциалов дает возможность проникновения К из эндолимфы внутрь волосковых клеток. Изменение положения одного волоска открывает 200-300 К- каналов и возникает деполяризация. Закрытие сопровождается гиперполяризацией. В кортиевом органе идет частотное кодирование за счет возбуждения разных участков основной мембраны. При этом было показано что звуки низкой частоты могут кодироваться числом нервных импульсов таким же количеством как и звуком. Такое кодирование возможно при восприятии звука до 500Гц. Кодирование информации звука достигается увеличением числа залпов волокон на более интенсивный звук и за счет числа активирующихся нервных волокон. Чувствительные волокна спирального ганглия оканичиваются в дорсальных и вентральных ядрах улитки продолговатого мозга. От этих ядер сигнал поступает в ядра оливы как своей так и противоположной стороны. От ее нейронов идут восходящие пути в составе латеральной петли которые подходят к нижним бугоркам четверохолмия и медиальному коленчатому телу зрительного бугра. От последнего сигнал идет в верхнюю височную извилину(извилина Гешля). Это соответствует 41 и 42 полям(первичная зона) и 22 поле(вторичная зона). В ЦНС существует топотоническая организация нейронов, то есть воспринимаются звуки с разной частотой и разной интенсивностью. Корковый центр имеет значение для восприятия, последовательности звука и пространственной локализации. При поражении 22 поля нарушается определение слов (рецептивная оппозия).
Ядра верхней оливы делят на медиальные и латеральные части. А латеральные ядра определяют неодинаковую интенсивность звуков, поступающих к обеим ушам. Медиальное ядро верхней оливы улавливает временные различия поступления звуковых сигналов. Обнаружено что сигналы от обоих ушей поступают в различные дендритные системы одного и того же воспринимающего нейрона. Нарушение слухового восприятия может проявляться звоном в ушах при раздражении внутреннего уха или слухового нерва и двумя типами глухоты: проводниковой и нервной. Первая связана с поражениями наружного и среднего уха(серная пробка).Вторая связана с дефектами внутреннего уха и поражениями слухового нерва. У пожилых людей утрачивается способность воспринимать высокочастотные голоса. За счет двух ушей можно определять пространственную локализацию звука. Это оказывается возможным, если звук отклоняется от средины положения на 3 градуса. При восприятии звуков возможно развитие адаптации за счет ретикулярной формации и эфферентных волокон(воздействием на наружные волосковые клетки.
Зрительная система.
Зрение – многозвеньевой процесс, начинающийся с проекции изображения на сетчатку глаза, затем идёт возбуждение фоторецепторов, передача и преобразование в нейронных слоях зрительной системы и заканчивается принятием высшими корковыми отделами решения о зрительном образе.
Строение и функции оптического аппарата глаза. Глаз имеет шарообразную форму, что важно для поворота глаза. Свет проходит через несколько прозрачных сред – роговицу, хрусталик и стекловидное тело, имеющие определённые преломляющие силы, выражающихся в диоптриях. Диоптрия равна преломляющей силе линзы с фокусным расстоянием 100 см. Преломляющая сила глаза при рассматривании далёких предметов – 59D, близких – 70,5D. На сетчатке образуется уменьшенное перевёрнутое изображение.
Аккомодация – приспособление глаза к ясному видению предметов на разных расстояниях. Хрусталик играет главную роль в аккомодации. При рассмотрении близких предметов ресничные мышцы сокращаются, циннова связка расслабляется, хрусталик становится более выпуклым в силу его эластичности. При рассмотрении дальних – мышцы расслаблены, связки натянуты и растягивают хрусталик, делая его более уплощённым. Ресничные мышцы иннервируются парасимпатическими волокнами глазодвигательного нерва. В норме дальняя точка ясного видения – в бесконечности, ближайшая – 10 см от глаза. Хрусталик с возрастом теряет эластичность, поэтому ближайшая точка ясного видения отодвигается и развивается старческая дальнозоркость.
Аномалии рефракции глаза.
Близорукость (миопия). Если продольная ось глаза слишком длинная или увеличивается преломляющая сила хрусталика, то изображение фокусируется перед сетчаткой. Человек плохо видит вдаль. Назначаются очки с вогнутыми стёклами.
Дальнозоркость (гиперметропия). Развивается при уменьшении преломляющих сред глаза или при укорочении продольной оси глаза. В результате изображение фокусируется за сетчаткой и чел плохо видит близкорасположенные предметы. Назначаются очки с выпуклыми линзами.
Астигматизм – неодинаковое преломление лучей в разных направлениях, обусловленное не строго сферической поверхностью роговой оболочки. Компенсируются очками с поверхностью, приближающейся к цилиндрической.
Зрачок и зрачковый рефлекс. Зрачок – отверстие в центре радужной оболочки, через которое лучи света проходят внутрь глаза. Зрачок повышает чёткость изображения на сетчатке, увеличивая глубину резкости глаза и за счёт устранения сферической аберрации. Если прикрыть глаз от света, а затем открыть его, то зрачок быстро сужается – зрачковый рефлекс. На ярком свету размер – 1,8 мм, при среднем – 2,4, в темноте – 7,5. Увеличение приводит к ухудшению качества изображения, но повышает чувствительность. Рефлекс имеет адаптационное значение. Расширяет зрачок симпатика, сужает – парасимпатика. У здоровых размеры обоих зрачков одинаковы.
Структура и функции сетчатки. Сетчатка – внутренняя светочувствительная оболочка глаза. Слои:
Пигментный – ряд отростчатых эпителиальных клеток чёрного цвета. Функции: экранирование (препятствует рассеиванию и отражению света, повышая чёткость), регенерация зрительного пигмента, фагоцитоз обломков палочек и колбочек, питание фоторецепторов. Контакт между рецепторами и пигментным слоем слабая, поэтому именно здесь происходит отслойка сетчатки.
Фоторецепторы. Колбы отвечают за цветовое зрение, их – 6-7 млн. Палки за сумеречное, их – 110-123 млн. Они расположены неравномерно. В центральной ямке – только колбы, здесь – наибольшая острота зрения. Палки чувствительнее колб.
Строение фоторецептора. Состоит из наружной воспринимающей части – наружного сегмента, с зрительным пигментом; соединительной ножки; ядерной части с пресинаптическим окончанием. Наружная часть состоит из дисков – двумембранная структура. Наружные сегменты постоянно обновляются. Пресинаптическое окончание содержит глутамат.
Зрительные пигменты. В палках – родопсин с поглощением в области 500 нм. В колбах – йодопсин с поглощениями 420 нм (синий), 531 нм (зелёный), 558 (красный). Молекула состоит из белка опсина и хромофорной части – ретиналя. Только цис-изомер воспринимает свет.
Физиология фоторецепции. При поглощении кванта света цис-ретиналь превращается в транс-ретиналь. Это вызывает пространственные изменения в белковой части пигмента. Пигмент обесцвечивается и переходит в метародопсин II, способный взаимодействовать с примембранным белком трансдуцином. Трансдуцин активируется и связывается с ГТФ, активируя фосфодиэстеразу. ФДЭ разрушает цГМФ. В результате концентрация цГМФ падает, что приводит к закрытию ионных каналов, при этом понижается концентрация натрия, приводя к гиперполяризации и возникновению рецепторного потенциала, распостраняющимся по клетке до пресинаптического окончания и вызывая уменьшение выделения глутамата.
Восстановление исходного темнового состояния рецептора. При утрате метародопсином способности взаимодействовать с трандуцином и активируется гуанилатциклаза, синтезирующая цГМФ. Гуанилатциклаза активируется падением концентрации кальция, выбрасываемого из клетки белком-обменником. В результате концентрация цГМФ повышается и она вновь связывается с ионным каналом, открывая его. При открытии в клетку идут натрий и кальций, деполяризуя мембрану рецептора, переводя его в темновое состояние, что вновь ускоряет выход медиатора.
Нейроны сетчатки.
Фоторецепторы синаптически связаны с биполярными нейронами. При действии света на медиатор уменьшается выделение медиатора, что приводит к гиперполяризации биполярного нейрона. От биполярного сигнал передаётся на ганглиозный. Импульсы от многих фоторецепторов конвергируют к одному ганглиозному нейрону. Взаимодействие соседних нейронов сетчатки обеспечивается горизонтальными и амакриновыми клетками, сигналы которых меняют синаптическую передачу межде рецепторами и биполярными (горизонтальные) и между биполярными и ганглиозными (амакриновые). Амакриновые клетки осуществляют боковое торможение между соседними ганглиозными клетками. В системе есть и эфферентные волокна, действующие на синапсы между биполярными и ганглиозными клетками, регулируя возбуждение меж ними.
Нервные пути.
1ый нейрон – биполярный.
2ой – ганглиозный. Их отростки идут в составе зрительного нерва, делают частичный перекрёст (необходимо для обеспечения каждого полушария информацией от каждого глаза) и идут в мозг в составе зрительного тракта, попадая в латеральное коленчатое тело таламуса (3ий нейрон). Из таламуса – в проекционную зону коры 17ое поле. Здесь 4ый нейрон.
Зрительные функции.
Абсолютная чувствительность. Для возникновения зрительного ощущения необходимо, чтобы световой раздражитель имел минимальную (пороговую) энергию. Палка может быть возбуждена одним квантом света. Палки и колбы мало различаются по возбудимости, но число рецепторов, посылающих сигналы на одну ганглиозную клетку различно в центре и на периферии.
Зрительная алаптация.
Приспособление зрительной сенсорной системы к условиям яркрй освещённости – световая адаптация. Обратное явление – темновая адаптация. Повышение чувствительности в темноте – поэтапное, обусловленное темновым восстановлением зрительных пигментов. Сначала восстанавливается йодопсин колб. Это мало влияет на чувствительность. Затем восстанавливается родопсин палок, что очень сильно повышает чувствительность. Для адаптации так же важны процессы изменения связей между элементами сетчатки: ослабление горизонтального торможения, приводящее к увеличению числа клеток, посылающее сигналы на ганглиозный нейрон. Влияние ЦНС тоже играет роль. При освещении одного глаза понижает чувствительность другого.
Дифференциальная зрительная чувствительность. По закону Вебера человек различит разницу в освещении, если оно будет сильнее на 1-1,5%.
Яркостной контраст происходит из-за взаимного латерального торможения зрительных нейронов. Серая полоска на светлом фоне кажется темнее серой на тёмном, так как клетки возбуждённые светлым фоном тормозят клетки, возбуждённые серой полоской.
Слепящая яркость света . Слишком яркий свет вызывает неприятное ощущение ослепления. Верхняя граница слепящей яркости зависит от адаптации глаза. Чем дольше была темновая адаптация, тем меньшая яркость вызывает ослепление.
Инерция зрения. Зрительное ощущение появляется и пропадает не сразу. От раздражения до восприятия проходит 0,03-0,1 с. Быстро следующие одно за другим раздражения сливаются в одно ощущение. Минимальная частота следования световых стимулов, при которой происходит слияние отдельных ощущений, называется критической частотой слития мельканий. На этом основано кино. Ощущения, продолжающиеся после прекращения раздражения – последовательные образы (образ лампы в темноте после её выключения).
Цветовое зрение.
Весь видимый спектр от фиолетового (400нм) до красного (700нм).
Теории. Трёхкомпонентная теория Гельмгольца. Цветовое ощущение обеспечиваемое тремя типами колб, чувствительных к одной части спектра (красной, зелёной или синей).
Теория Геринга. В колбах есть вещества чувствительные к бело-чёрному, красно-зелёному и жёлто-синему излучениям.
Последовательные цветовые образы. Если смотреть на окрашенный предмет, а затем на белый фон, то фон приобретёт дополнительный цвет. Причина – цветовая адаптация.
Цветовая слепота. Дальтонизм – расстройство, при котором невозможно различие цветов. При протанопии не различается красный цвет. При дейтеранопии – зелёный. При тританопии – синий. Диагностируется полихроматическими таблицами.
Полная потеря цветовосприятия – ахромазия, при которой всё видится в оттенках серого.
2.Классификация и свойства рецепторов. Механизмы возбуждения первично- и вторичночувствующих рецепторов.
Большинство рецепторов возбуждаются в ответ на действие стимулов только одной физической природы и поэтому относятся к мономодальным. Их можно возбудить и некоторыми неадекватными раздражителями , например фоторецепторы - сильным давлением на глазное яблоко, а вкусовые рецепторы - прикосновением языка к контактам гальванической батареи, но получить качественно различаемые ощущения в таких случаях невозможно. Наряду с мономодальными существуют полимодальные рецепторы, адекватными стимулами которых могут служить раздражители разной природы. К такому типу рецепторов принадлежат некоторые болевые рецепторы, или ноцицепторы (лат. nocens - вредный), которые можно возбудить механическими, термическими и химическими стимулами. Полимодальность имеется у терморецепторов, реагирующих на повышение концентрации калия во внеклеточном пространстве так же, как на повышение температуры.
В зависимости от строения рецепторов их подразделяют на первичные
, или первичночувствующие
, которые являются специализированными окончаниями чувствительного нейрона, и вторичные
, или вторичночувствующие
, представляющие собой клетки эпителиального происхождения, способные к образованию рецепторного потенциала в ответ на действие адекватного стимула. Первичночувствующие рецепторы
могут сами генерировать потенциалы действия в ответ на раздражение адекватным стимулом, если величина их рецепторного потенциала достигнет пороговой величины. К ним относятся обонятельные рецепторы, большинство механорецепторов кожи, терморецепторы, болевые рецепторы
или ноцицепторы
, проприоцепторы и большинство интерорецепторов внутренних органов.
Вторичночувствующие рецепторы
отвечают на действие раздражителя лишь возникновением рецепторного потенциала
, от величины которого зависит количество выделяемого этими клетками медиатора. С его помощью вторичные рецепторы действуют на нервные окончания чувствительных нейронов, генерирующих потенциалы действия в зависимости от количества медиатора, выделившегося из вторичночувствующих рецепторов. Вторичные рецепторы
представлены вкусовыми, слуховыми и вестибулярными рецепторами, а также хемочувствительными клетками синокаротидного клубочка. Фоторецепторы сетчатки, имеющие общее происхождение с нервными клетками , чаще относят к первичным рецепторам, но отсутствие у них способности генерировать потенциалы действия указывает на их сходство с вторичными рецепторами.
В зависимости от источника адекватных стимулов рецепторы
подразделяют на наружные и внутренние, или экстерорецепторы
и интерорецепторы
; первые стимулируются при действии раздражителей внешней среды (электромагнитные и звуковые волны, давление, действие пахучих молекул), а вторые - внутренней (к этому типу рецепторов относят не только висцерорецепторы внутренних органов, но также проприоцепторы и вестибулярные рецепторы). В зависимости от того, действует стимул на расстоянии или непосредственно на рецепторы, их подразделяют еще на дистантные и контактные.
Классификация рецепторов. В основу классификации рецепторов положено несколько критериев.
Психофизиологический характер ощущения: тепловые, холодовые, болевые и др.
Природа адекватного раздражителя: механо-, термо-, хемо-, фото-, баро-, осмбрецепторы и др.
Среда, в которой рецептор воспринимает раздражитель: экстеро-, интерорецепторы.
Отношение к одной или нескольким модальностям: моно- и полимодальные (мономодальные преобразуют в нервный импульс только один вид раздражителя - световой, температурный и т. д., полимодальные могут несколько раздражителей преобразовать в нервный импульс - механический и температурный, механический и химический и т. д.).
Способность воспринимать раздражитель, находящийся на расстоянии от рецептора или при непосредственном контакте с ним: контактные и дистантные.
Уровень чувствительности (порог раздражения): низкопороговые (механорецепторы) и высокопороговые (ноцицепторы).
Скорость адаптации: быстроадаптирующиеся, (тактильные), медленноадаптирующиеся (болевые) и неадаптирующиеся (вестибулярные рецепторы и проприорецепторы).
Отношение к различным моментам действия раздражителя: при включении раздражителя, при его выключении, на протяжении всего времени действия раздражителя.
Морфофункциональная организация и механизм возникновения возбуждения: первичночувствующие и вторичночувствующие.
3.Вкусовая система: рецепторный, проводниковый и корковый отделы. Механизмы восприятия вкуса. Методы исследования вкусовой системы.
| Вкусовая система В процессе эволюции вкус формировался как механизм выбора или отвергания пищи. В естественных условиях вкусовые ощущения комбинируются с обонятельными, тактильными и термическими, также создаваемыми пищей. Важным обстоятельством является то, что предпочтительный выбор пищи отчасти основан на врожденных механизмах, но в значительной мере зависит от связей, выработанных в онтогенезе условнорефлекторным путем. Вкус, так же как и обоняние, основан на хеморецепции. Вкусовые рецепторы несут информацию о характере и концентрации веществ, поступающих в рот. Их возбуждение запускает сложную цепь реакций разных отделов мозга, приводящих к различной работе органов пищеварения или к удалению вредных для организма веществ, попавших в рот с пищей. Рецепторы вкуса. Вкусовые почки - рецепторы вкуса - расположены на языке, задней стенке глотки, мягком небе, миндалинах и надгортаннике. Больше всего их на кончике , краях и задней части языка. Каждая из примерно 10 000 вкусовых почек человека состоит из нескольких (2-6) рецепторных клеток и, кроме того, из опорных клеток. Вкусовая почка имеет колбовидную форму; у человека ее длина и ширина около 70 мкм. Вкусовая почка не достигает поверхности слизистой оболочки языка и соединена с полостью рта через вкусовую пору. Вкусовые клетки - наиболее короткоживущие эпителиальные клетки организма: в среднем через каждые 250 ч старая клетка сменяется молодой, движущейся к центру вкусовой почки от ее периферии. Каждая из рецепторных вкусовых клеток длиной 10- 20 мкм и шириной 3-4 мкм имеет на конце, обращенном в просвет поры, 30-40 тончайших микроворсинок толщиной 0,1- 0,2 мкм и длиной 1-2 мкм. Считают, что они играют важную роль в возбуждении рецепторной клетки, воспринимая те или иные химические вещества, адсорбированные в канале почки. Предполагают, что в области микроворсинок расположены активные центры - стереоспецифические участки рецептора, избирательно воспринимающие разные адсорбированные вещества. Этапы первичного преобразования химической энергии вкусовых веществ в энергию нервного возбуждения вкусовых рецепторов еще не известны. Электрические потенциалы вкусовой системы. В опытах с введением микроэлектрода внутрь вкусовой почки животных показано, что суммарный потенциал рецепторных клеток изменяется при раздражении языка разными веществами (сахар, соль, кислота). Этот потенциал развивается довольно медленно: максимум его достигается к 10-15-й секунде после воздействия, хотя электрическая активность в волокнах вкусового нерва начинается значительно раньше. Проводящие пути и центры вкуса. Проводниками всех видов вкусовой чувствительности служат барабанная струна и языкоглоточный нерв, ядра которых в продолговатом мозге содержат первые нейроны вкусовой системы. Многие из волокон, идущих от вкусовых рецепторов, отличаются определенной специфичностью, так как отвечают учащением импульсных разрядов лишь на действие соли, кислоты и хинина. Другие волокна реагируют на сахар. Наиболее убедительной считается гипотеза, согласно которой информация о 4 основных вкусовых ощущениях: горьком, сладком, кислом и соленом - кодируется не импульсацией в одиночных волокнах, а разным распределением частоты разрядов в большой группе волокон, по-разному возбуждаемых вкусовым веществом. Вкусовые афферентные сигналы поступают в ядро одиночного пучка ствола мозга. От ядра одиночного пучка аксоны вторых нейронов восходят в составе медиальной петли до дугообразного ядра таламуса , где расположены третьи нейроны, аксоны которых направляются в корковый центр вкуса. Результаты исследований пока не позволяют оценить характер преобразований вкусовых афферентных сигналов на всех уровнях вкусовой системы. Вкусовые ощущения и восприятие. У разных людей абсолютные пороги вкусовой чувствительности к разным веществам существенно отличаются вплоть до «вкусовой слепоты» к отдельным агентам (например, к креатину). Абсолютные пороги вкусовой чувствительности во многом зависят от состояния организма (они изменяются в случае голодания, беременности и т.д.). При измерении абсолютной вкусовой чувствительности возможны две ее оценки: возникновение неопределенного вкусового ощущения (отличающегося от вкуса дистиллированной воды) и осознанное восприятие или опознание определенного вкуса. Порог восприятия, как и в других сенсорных системах, выше порога ощущения. Пороги различения минимальны в диапазоне средних концентраций веществ, но при переходе к большим концентрациям резко повышаются. Поэтому 20 % раствор сахара воспринимается как максимально сладкий, 10 % раствор натрия хлорида - как максимально соленый, 0,2 % раствор соляной кислоты - как максимально кислый, а 0,1 % раствор хинина сульфата - как максимально горький. Пороговый контраст (dI/I) для разных веществ значительно колеблется. Вкусовая адаптация. При длительном действии вкусового вещества наблюдается адаптация к нему (снижается интенсивность вкусового ощущения). Продолжительность адаптации пропорциональна концентрации раствора. Адаптация к сладкому и соленому развивается быстрее, чем к горькому и кислому. Обнаружена и перекрестная адаптация, т. е. изменение чувствительности к одному веществу при действии другого. Применение нескольких вкусовых раздражителей одновременно или последовательно дает эффекты вкусового контраста или смешения вкуса. Например, адаптация к горькому. повышает чувствительность к кислому и соленому, адаптация к сладкому обостряет восприятие всех других вкусовых стимулов. При смешении нескольких вкусовых веществ может возникнуть новое вкусовое ощущение, отличающееся от вкуса составляющих смесь компонентов. |
4.Обонятельная система: рецепторы, их локализация, механизм восприятия запахов, проводниковый и корковый отделы.
Обонятельная система
Обонятельная рецепторная клетка - биполярная клетка, на апикальном полюсе которой находятся реснички, а от ее базальной части отходит немиелинизированный аксон. Аксоны рецепторов образуют обонятельный нерв, который пронизывает основание черепа и вступает в обонятельную луковицу. Подобно вкусовым клеткам и наружным сегментам фоторецепторов, обонятельные клетки постоянно обновляются. Продолжительность жизни обонятельной клетки около 2 мес.
Молекулы пахучих веществ попадают в слизь, вырабатываемую обонятельными железами, с постоянным током воздуха или из ротовой полости во время еды. Принюхивание ускоряет приток пахучих веществ к слизи. В слизи молекулы пахучих веществ на короткое время связываются с обонятельными нерецепторными белками. Некоторые молекулы достигают ресничек обонятельного рецептора и взаимодействуют с находящимся в них обонятельным рецепторным белком. В свою очередь обонятельный белок активирует , как и в случае фоторецепции, ГТФ-связывающий белок (G-белок), а тот в свою очередь - фермент аденилатциклазу, синтезирующую цАМФ. Повышение в цитоплазме концентрации цАМФ вызывает открывание в плазматической мембране рецепторной клетки натриевых каналов и как следствие - генерацию деполяризационного рецепторного потенциала. Это приводит к импульсному разряду в аксоне рецептора (волокне обонятельного нерва).
Раньше считали, что низкая избирательность отдельного рецептора объясняется наличием в нем множества типов обонятельных рецепторных белков, однако недавно выяснено, что каждая обонятельная клетка имеет только один тип мембранного рецепторного белка. Сам же этот белок способен связывать множество пахучих молекул различной пространственной конфигурации. Правило «одна обонятельная клетка - один обонятельный рецепторный белок» значительно упрощает передачу и обработку информации о запахах в обонятельной луковице - первом нервном центре переключения и обработки хемосенсорной информации в мозге.
Наличие всего одного обонятельного белка в каждом рецепторе обусловлено не только тем, что каждая обонятельная клетка эк-спрессирует только один из сотен генов обонятельных белков, но и тем, что в пределах данного гена экспрессируется только одна из двух аллелей - материнская или отцовская. Вероятно, что генетически обусловленные индивидуальные различия в порогах восприятия определенных запахов связаны с функциональными отличиями в механизмах экспрессии гена обонятельного рецепторного белка.
Электроольфактограмма. Суммарный электрический потенциал, регистрируемый от поверхности обонятельного эпителия, называют электроольфактограммой (рис. 14.24). Это монофазная негативная волна с амплитудой до 10 мВ и длительностью несколько секунд, возникающая в обонятельном эпителии даже при кратковременном воздействии на него пахучего вещества. Нередко на электроольфактограмме можно видеть небольшое позитивное отклонение потенциала, предшествующее основной негативной волне, а при достаточной длительности воздействия регистрируется большая негативная волна на его прекращение (off-реакция). Иногда на медленные волны электроольфактограммы накладываются быстрые осцилляции, отражающие синхронные импульсные разряды значительного числа рецепторов.
Кодирование обонятельной информации. Как показывают исследования с помощью микроэлектродов, одиночные рецепторы отвечают увеличением частоты импульсации, которое зависит от качества и интенсивности стимула. Каждый обонятельный рецептор отвечает не на один, а на многие пахучие вещества, отдавая «предпочтение» некоторым из них. Считают, что на этих свойствах рецепторов , различающихся по своей настройке на разные группы веществ, может быть основано кодирование запахов и их опознание в центрах обонятельной сенсорной системы. При электрофизиологических исследованиях обонятельной луковицы выявлено, что регистрируемый в ней при действии запаха электрический ответ зависит от пахучего вещества: при разных запахах меняется пространственная мозаика возбужденных и заторможенных участков луковицы. Служит ли это способом кодирования обонятельной информации, пока судить трудно.
Центральные проекции обонятельной системы. Особенность обонятельной системы состоит, в частности, в том, что ее афферентные волокна не переключаются в таламусе и не переходят на противоположную сторону большого мозга. Выходящий из луковицы обонятельный тракт состоит из нескольких пучков, которые направляются в разные отделы переднего мозга: переднее обонятельное ядро, обонятельный бугорок, препириформную кору, периамигдалярную кору и часть ядер миндалевидного комплекса. Связь обонятельной луковицы с гиппокампом, пириформной корой и другими отделами обонятельного мозга осуществляется через несколько переключений. Показано, что наличие значительного числа центров обонятельного мозга (rhinencephalon) не является необходимым для опознания запахов, поэтому большинство нервных центров, в которые проецируется обонятельный тракт, можно рассматривать как ассоциативные центры, обеспечивающие связь обонятельной сенсорной системы с другими сенсорными системами и организацию на этой основе ряда сложных форм поведения - пищевой, оборонительной, половой и т. д.
Эфферентная регуляция активности обонятельной луковицы изучена пока недостаточно, хотя есть морфологические предпосылки, свидетельствующие о возможности таких влияний.
Чувствительность обонятельной системы человека. Эта чувствительность чрезвычайно велика: один обонятельный рецептор может быть возбужден одной молекулой пахучего вещества, а возбуждение небольшого числа рецепторов приводит к возникновению ощущения. В то же время изменение интенсивности действия веществ (порог различения) оценивается людьми довольно грубо (наименьшее воспринимаемое различие в силе запаха составляет 30-60 % от его исходной концентрации). У собак эти показатели в 3-6 раз выше. Адаптация в обонятельной системе происходит сравнительно медленно (десятки секунд или минуты) и зависит от скорости потока воздуха над обонятельным эпителием и от концентрации пахучего вещества.
5.Вестибулярная система и ее роль в оценке положения тела в пространстве и при движении. Рецепторный, проводниковый и корковый отделы. Результаты одно- и двухстороннего разрушения лабиринтов у лягушки.
Вестибулярная система
Вестибулярная система играет наряду со зрительной и соматосенсорной системами ведущую роль в пространственной ориентировке человека. Она получает, передает и анализирует информацию об ускорениях или замедлениях, возникающих в процессе прямолинейного или вращательного движения, а также при изменении положения головы в пространстве. При равномерном движении или в условиях покоя рецепторы вестибулярной сенсорной системы не возбуждаются. Импульсы от вестибулорецепторов вызывают перераспределение тонуса скелетной мускулатуры , что обеспечивает сохранение равновесия тела. Эти влияния осуществляются рефлекторным путем через ряд отделов ЦНС.
Строение и функции рецепторов вестибулярной системы . Периферическим отделом вестибулярной системы является вестибулярный аппарат, расположенный в лабиринте пирамиды височной кости. Он состоит из преддверия (vestibulum) и трех полукружных каналов (canales cemicircularis). Кроме вестибулярного аппарата, в лабиринт входит улитка, в которой располагаются слуховые рецепторы. Полукружные каналы (рис. 14.17) располагаются в трех взаимно перпендикулярных плоскостях: верхний - во фронтальной, задний - в сагиттальной и латеральный - в горизонтальной. Один из концов каждого канала расширен (ампула).
Вестибулярный аппарат включает в себя также два мешочка: сферический (sacculus) и эллиптический, или маточку (utriculus). Первый из них лежит ближе к улитке, а второй - к полукружным каналам. В мешочках преддверия находится отолитовый аппарат: скопления рецепторных клеток (вторично-чувствующие механорецепторы) на возвышениях, или пятнах (macula sacculi, macula utriculi). Выступающая в полость мешочка часть рецепторной клетки оканчивается одним более длинным подвижным волоском и 60-80 склеенными неподвижными волосками. Эти волоски пронизывают желеобразную мембрану, содержащую кристаллики карбоната кальция - отолиты. Возбуждение волосковых клеток преддверия происходит вследствие скольжения отоли-товой мембраны по волоскам, т. е. их сгибания (рис. 14.18).
В перепончатых полукружных каналах, заполненных, как и весь лабиринт, плотной эндолимфой (ее вязкость в 2-3 раза больше, чем у воды), рецепторные волосковые клетки сконцентрированы только в ампулах в виде крист (cristae ampularis). Они также снабжены волосками. При движении эндолимфы (во время угловых ускорений), когда волоски сгибаются в одну сторону, волосковые клетки возбуждаются, а при противоположно направленном движении - тормозятся. Это связано с тем, что механическое управление ионными каналами мембраны волоска с помощью микрофиламентов, описанное в разделе «механизмы слуховой рецепции», зависит от направления сгиба волоска: отклонение в одну сторону приводит к открыванию каналов и деполяризации волосковой клетки, а отклонение в противоположном направлении вызывает закрытие каналов и гиперполяризацию рецептора. В волосковых клетках преддверия и ампулы при их сгибании генерируется рецепторный потенциал, который усиливает выделение ацетилхолина и через синапсы активирует окончания волокон вестибулярного нерва.
Волокна вестибулярного нерва (отростки биполярных нейронов) направляются в продолговатый мозг. Импульсы, приходящие по этим волокнам, активируют нейроны бульбарного вестибулярного комплекса, в состав которого входят ядра: преддверное верхнее, или Бехтерева, преддверное латеральное, или Дейтерса, Швальбе и др. Отсюда сигналы направляются во многие отделы ЦНС: спинной мозг, мозжечок, глазодвигательные ядра, кору большого мозга , ретикулярную формацию и ганглии автономной нервной системы.
Комплексные рефлексы, связанные с вестибулярной стимуляцией. Нейроны вестибулярных ядер обеспечивают контроль и управление различными двигательными реакциями. Важнейшими из этих реакций являются следующие: вестибулоспинальные, вестибуловегетативные и вестибулоглазодвигательные. Вестибулоспинальные влияния через вестибуло-, ретикуло- и руброспинальные тракты изменяют импульсацию нейронов сегментарных уровней спинного мозга. Так осуществляется динамическое перераспределение тонуса скелетной мускулатуры и включаются рефлекторные реакции, необходимые для сохранения равновесия. Мозжечок при этом ответствен за фазический характер этих реакций: после его удаления вестибулоспинальные влияния становятся по преимуществу тоническими. Во время произвольных движений вестибулярные влияния на спинной мозг ослабляются.
Вестибулоглазодвигательные рефлексы (глазной нистагм) состоят в медленном движении глаз в противоположную вращению сторону, сменяющемся скачком глаз обратно. Само возникновение и характеристика вращательного глазного нистагма - важные показатели состояния вестибулярной системы, они широко используются в морской, авиационной и космической медицине, а также в эксперименте и клинике.
Основные афферентные пути и проекции вестибулярных сигналов. Есть два основных пути поступления вестибулярных сигналов в кору большого мозга: прямой - через дорсомедиальную часть вентрального постлатерального ядра и непрямой вестибулоцеребеллоталамический путь через медиальную часть вентролатерального ядра. В коре полушарий большого мозга основные афферентные проекции вестибулярного аппарата локализованы в задней части постцентральной извилины. В моторной зоне коры спереди от нижней части центральной борозды обнаружена вторая вестибулярная зона.
Функции вестибулярной системы. Вестибулярная система помогает организму ориентироваться в пространстве при активном и пассивном движении.
При пассивном движении корковые отделы системы запоминают направление движения, повороты и пройденное расстояние. Следует подчеркнуть, что в нормальных условиях пространственная ориентировка обеспечивается совместной деятельностью зрительной и вестибулярной систем. Чувствительность вестибулярной системы здорового человека очень высока: отолитовый аппарат позволяет воспринять ускорение прямолинейного движения, равное всего 2 см/с2. Порог различения наклона головы в сторону - всего около 1°, а вперед и назад - 1,5-2°. Рецепторная система полукружных каналов позволяет человеку замечать ускорения вращения 2-3°∙ с-2.