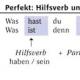
Енергетичний обмін та загальний шлях катаболізму. Біохімічні шляхи пристосовності організму до умов геохімічного середовища До основних функцій мембран можна віднести
Зміст статті
БІОХІМІЯ,наука, яка описує мовою хімії будову та функції живих організмів. Біохімічні концепції знаходять застосування в медицині, харчовій, фармацевтичній та мікробіологічній промисловості, сільському господарстві, а також у переробній промисловості, яка використовує відходи та побічні продукти сільського господарства.
Області досліджень.
У розвитку біохімії можна назвати кілька етапів і напрямів.
Типи органічних сполук та його структура.
Фундаментальне значення мало складання переліку органічних сполук, виявлених у живих організмах, та встановлення структури кожного з них. Цей перелік включає відносно прості сполуки – амінокислоти, цукру та жирні кислоти, потім складніші – пігменти (надають забарвлення, наприклад, квіткам), вітаміни та коферменти (небілкові компоненти ферментів), а закінчується гігантськими молекулами білків та нуклеїнових кислот.
Метаболічні шляхи.
Очевидно, найбільш значні успіхи у біохімії пов'язані з з'ясуванням шляхів біосинтезу природних сполук із найпростіших речовин, тобто. з компонентів їжі у тварин та з діоксиду вуглецю та мінеральних речовин (під час фотосинтезу) у рослин. Біохімікам вдалося докладно вивчити основні метаболічні шляхи, що забезпечують синтез та розщеплення природних сполук у тварин, рослин та мікроорганізмів (зокрема, у бактерій).
Структура та функції макромолекул.
Третій напрямок біохімії пов'язаний з аналізом зв'язку між структурою та функцією біологічних макромолекул. Так, біохіміки намагаються зрозуміти, які особливості структури білкових каталізаторів є основою їх специфічності, тобто. можливості прискорювати строго певні реакції; як виконують свої функції складні полісахариди, що входять до складу клітинних стінок та мембран; як складні ліпіди, присутні у нервової тканини, беруть участь у функціонуванні нервових клітин – нейронів.
Функціонування клітин.
Ще одна проблема, якою займаються біохіміки – розкриття механізмів функціонування спеціалізованих клітин. Досліджуються, наприклад, такі питання: як відбувається скорочення м'язових клітин, як певні клітини формують кісткову тканину, яким чином еритроцити переносять кисень від легень до тканин і забирають із тканин вуглекислий газ, який механізм синтезу пігментів у клітинах рослин тощо.
Генетичні аспекти
Дослідження, що почалися в 1940-х роках і проводилися на грибах і бактеріях, а потім на вищих організмах, включаючи людину, показали, що зазвичай внаслідок мутації генів у клітинах перестають протікати певні біохімічні реакції. Ці спостереження спричинили створення концепції гена як інформаційної одиниці, що відповідає за синтез специфічного білка. Якщо білок є ферментом, а кодує його ген зазнав мутації (тобто змінився), то клітина втрачає здатність здійснювати реакцію, яку цей фермент мав би каталізувати.
Ген – це специфічний сегмент молекули дезоксирибонуклеїнової кислоти (ДНК), який здатний реплікуватися (відтворювати себе) та відповідальний за синтез певного білка. Багато біохімічних досліджень спрямовані на з'ясування деталей реплікації нуклеїнових кислот та механізму синтезу білків, а тому тісно пов'язані з генетикою. Область досліджень, що лежить у сфері та біохімії та генетики, зазвичай називають молекулярною біологією.
Проект «Геном людини»
– грандіозний міжнародний проект у галузі молекулярної біології та генетики, у якому беруть участь колективи вчених із багатьох країн. Мета проекту – побудувати генетичні карти 23 хромосом людини із точним вказівкою становища всіх десятків тисяч генів цих хромосомах й у остаточному підсумку визначити структуру хромосом, тобто. послідовність приблизно 3 млрд. пар азотистих основ, у тому числі складається хромосомна ДНК. Ці дослідження дозволять створити доступну всім вчених базу даних, які мають велику цінність вивчення генетики людини, а головне – допоможуть біохімікам розкрити механізми спадкових хвороб.
Медична біохімія.
З кожним роком дедалі більше хвороб вдається пов'язати з тими чи іншими порушеннями метаболізму. Спільні зусилля біохіміків та лікарів дозволили розкрити природу порушень, що лежать в основі таких захворювань, як цукровий діабет та серповидноклітинна анемія. Більш ніж у 800 випадках встановлено кореляцію між порушеннями метаболізму та генетичними дефектами, у деяких випадках знайдено способи, що дозволяють пом'якшити наслідки захворювання.
ТЕМА 6. ПЕРЕВОРЮВАННЯ ВУГЛІВНИКІВ У ТРАВЛІЧНОМУ ТРАКТІ. Гліколіз. ОКИСЛЮВАЛЬНЕ ДЕКАРБОКСИЛЮВАННЯ ПІРУВАТА
ЧАСТИНА 2. ДИНАМІЧНА БІОХІМІЯ
Гормони тимусу (вилочкова залоза)
Гормони паращитовидної залози
Гормони статевих залоз
Чоловічі статеві гормони ( андрогени) утворюються в сім'яниках, жіночі статеві гормони ( естрогени,прогестини) продукуються переважно у яєчниках. Статеві гормони є похідними циклопентапергідрофенантрену.
Андрогенистимулюють розвиток ембріона за чоловічим типом, відповідальні за формування вторинних статевих ознак (будова тіла, волосяний покрив, тембр голосу, процес сперматогенезу), мають виражений анаболічний ефект (стимулюють білковий синтез у скелетній мускулатурі). Найбільш важливим андрогеном є тестостерон. Похідні тестостерону мають ще сильнішу анаболічну дію. Побічна дія цих препаратів, що загрожує здоров'ю спортсменів, стала причиною заборони їх використання у спорті.
Біологічна роль естрогеніві прогестинівполягає у забезпеченні репродуктивної функції: оптимальних умов запліднення яйцеклітини, вагітності, пологів та ін. Основними представниками жіночих статевих гормонів є естрадіолі прогестерон.
Паращитовидні залози секретують 2 гормони ( паратгормоні кальцитонін), які разом із вітаміном Д забезпечують регуляцію кальцієвого обміну.
Паратгормонмає поліпептидну природу. Секреція паратгормону залежить від концентрації іонізованого кальцію в плазмі: підвищення концентрації кальцію знижує секрецію гормону, зниження концентрації кальцію – підвищує. Паратгормон збільшує канальцеву реабсорбцію кальцію та магнію, знижує зворотну реабсорбцію калію та фосфату, збільшує всмоктування кальцію у шлунково-кишковому тракті, мобілізує вихід кальцію з кісткової тканини в кров.
Кальцітонін– антогоніст паратгормону, кальцитонін зменшує розсмоктування кісткової тканини, зменшує екскрецію кальцію із сечею. Свій ефект кальцитонін здійснює через регуляцію роботи кальцієвого насоса.
У тимусі продукується 5 гормонально-активних факторів: тимозин, гомеостатичний тимусний гормон, тимопоетини 1і 11 , тимусний гуморальний фактор. Усі вони є поліпептидами. Основна функція гормонів вилочкової залози – регулювання дозрівання певних популяцій лімфоїдних клітин, тобто участь у регуляції функціонування імунної системи.
Характерна риса живих організмів – здатність до обміну речовин, що здійснює безліч різноманітних хімічних реакцій. Тварини організми отримують із довкілля поживні речовини і кисень і повертають у ній кінцеві продукти обміну – вуглекислий газ, воду, аміак, сечовину та інших. Життя тварин характеризується безперервним обміном речовин. Хімічні реакції в організмі узгоджені у часі та у просторі та протікають у певній послідовності. Процеси обміну речовин регулюються ЦНС та ендокринною системою. Обмін речовин здійснюється з допомогою біологічних каталізаторів – ферментів, які забезпечують закономірність хімічних процесів. Виключення будь-якого ферменту призводить до порушення нормального перебігу обміну речовин, що призводить до певної патології. В обміні речовин виділяють зовнішнійобмін та проміжний. Зовнішнійобмін - позаклітинне перетравлення речовин на шляхах їх надходження та виділення з організму. Під проміжним обміномрозуміють всю сукупність ферментативних реакцій, які у клітині.
Метаболізм виконує 4 основні функції: 1) вилучення енергії з довкілля (або у формі хімічної енергії органічних речовин або у формі енергії сонячного світла); 2) перетворення екзогенних речовин на будівельні блоки – на попередники макромолекулярних компонентів клітини; 3) складання білків, нуклеїнових кислот, жирів та ін клітинних компонентів з цих будівельних блоків; 4) синтез та руйнування тих біомолекул, які необхідні для виконання різних специфічних функцій цієї клітини.
Послідовності метаболічних реакцій подібні у всіх живих форм, особливо у тій частині, що становить центральні метаболічні шляхи. Метаболічний шлях – це послідовність реакцій, які призводять до сукупності до певного продукту.
А → Б → В → Г → Д, де А – вихідна речовина (попередник), Б, В, Г – інтермедіати, Д – кінцевий продукт.
Кожен етап каталізується спеціальним ферментом. У більшості випадків метаболічні шляхи лінійні, але можуть бути циклічні. Циклічні шляхи зазвичай мають відгалуження, в яких якісь продукти виходять з ланцюга або входять до нього. Метаболічні шляхи поділяються на центральні та спеціальні. Центральні шляхи – загальні для розпаду та синтезу основних макромолекул вуглеводів, ліпідів, білків, нуклеїнових кислот. Ці шляхи подібні до будь-яких представників живого світу. Спеціальні шляхи – шляхи для синтезу та розпаду індивідуальних, спеціальних речовин, що потрібні в менших кількостях (гормони, кофактори та ін.). Метаболічні шляхи поділяють на 3 категорії – катаболічні, анаболічніі амфіболічні. Катаболічнішляхи - це ферментативне розщеплення порівняно великих харчових молекул - вуглеводів, жирів і білків - що здійснюється переважно за рахунок реакцій окислення (джерелом поживних речовин служить або навколишнє середовище, або внутрішньоклітинні депо - запаси, що раніше відклалися в самій клітині). У ході окислення великі молекули розщеплюються з утворенням дрібніших. Катаболізм супроводжується виділенням вільної енергії, укладеної у складних структурах великих органічних молекул, та запасанням її у формі енергії фосфатних зв'язків аденозинтрифосфату (АТФ).
Катаболізм включає 3 основні етапи. на 1 етапівеликі харчові молекули розщеплюються на їх складові будівельні блоки(Амінокислоти, моносахариди, жирні кислоти та ін.). на 2 етапівелика кількість продуктів, що утворилися на 1 стадії, перетворюються на простіші молекули, кількість яких невелика - ацетил-КоАта ін. Потім на 3 етапіці продукти окислюються до СО 2і води.
Анаболічні шляхи- Це ферментативний синтез порівняно великих клітинних компонентів із простих попередників. Оскільки процеси синтезу ведуть до збільшення розмірів молекул і до ускладнення їх структури, а це означає зменшення ентропії, ці процеси пов'язані зі споживанням вільної енергії, яка поставляється у формі енергії фосфатних зв'язків АТФ. Процес анаболізму включає також 3 стадії, в результаті чого утворюються біополімери.
Амфіболічнішляхи – двоїсті, пов'язують катаболічні та анаболічні шляхи.
Катаболізм та анаболізм протікають у клітинах одночасно, але часто локалізуються у різних ділянках клітини. При катаболізмі харчових речовин енергія, що витягується з них, запасається у формі енергії фосфатних зв'язків (АТФ). Енергія, необхідна анаболізму, вивільняється при дефосфорилировании АТФ. Таким чином, АТФ виконує функцію переносника хімічної енергії. Хімічна енергія може передаватися від катаболічних шляхів до анаболічних також за участю коферментів (головним чином, НАДФН).
Регуляція метаболізму може здійснюватися різних рівнях. Вона може ґрунтуватися на використанні основних властивостей ферментів, на активності регуляторних або алостеричних ферментів, на генетичній репресії та дерепресії синтезу ферментів або на дії ферментів.
Введення у метаболізм (біохімія)
Обмін речовин або метаболізм – це сукупність хімічних реакцій в організмі, які забезпечують його речовинами та енергією, необхідними для життєдіяльності. Процес метаболізму, що супроводжується утворенням більш простих сполук із складних, позначають терміном – катаболізм. Процес, що йде у зворотному напрямку і приводить, зрештою, до утворення складного продукту з більш простих - анаболізм. Анаболічні процеси супроводжуються споживанням енергії, катаболічні вивільненням.
Анаболізм та катаболізм не є простим зверненням реакцій. Анаболічні шляхи повинні відрізнятись від шляхів катаболізму хоча б однієї з ферментативних реакцій, щоб регулюватися незалежно, і за рахунок контролю активності цих ферментів регулюється сумарна швидкість розпаду та синтезу речовин. Ферменти, які визначають швидкість всього процесу загалом, називаються ключовими.
Більше того, шлях яким йде катаболізм тієї чи іншої молекули, може бути непридатним для її синтезу з енергетичних міркувань. Наприклад, розщеплення глюкози до пірувату, що протікають у печінці, являє собою процес, що складається з 11 послідовних стадій, що каталізуються специфічними ферментами. Здавалося б, синтез глюкози з пірувату може бути простим зверненням всіх цих ферментативних стадій її розпаду. Такий шлях видається на перший погляд і найприроднішим, і найбільш економічним. Однак насправді біосинтез глюкози (глюконеогенез) у печінці протікає інакше. Він включає лише 8 з 11 ферментативних стадій, що у її розпаді, а 3 відсутні стадії замінені у ньому зовсім іншим набором ферментативних реакцій, властивим лише цьому біосинтетичному шляху. Крім того, реакції катаболізму та анаболізму часто розділені мембранами та протікають у різних компартментах клітин.
Таблиця 8.1. Компартменталізація деяких метаболічних шляхів у гепатоциті
|
Компартмент |
Метаболічні шляхи |
|---|---|
|
Цитозоль |
Гліколіз, багато реакцій глюконеогенезу, активація амінокислот, синтез жирних кислот |
|
Плазматична мембрана |
Енергозалежні транспортні системи |
|
Реплікація ДНК, синтез різних видів РНК |
|
|
Рибосоми |
Синтез білка |
|
Лізосоми |
Ізоляція гідролітичних ферментів |
|
Комплекс Гольджі |
Утворення плазматичної мембрани та секреторних бульбашок |
|
Мікросоми |
Локалізація каталази та оксидаз амінокислот |
|
Ендоплазматична мережа |
Синтез ліпідів |
|
Мітохондрії |
Цикл трикарбонових кислот, ланцюг тканинного дихання, окиснення жирних кислот, окисне фосфорилювання |
Метаболізм виконує 4 функції:
1. постачання організму хімічною енергією, отриманої при розщепленні багатих на енергію харчових речовин;
2. перетворення харчових речовин на будівельні блоки, які використовуються в клітині для біосинтезу макромолекул;
3. складання макромолекулярних (біополімери) та надмолекулярних структур живого організму, пластична та енергетична підтримка його структури;
4. синтез та руйнування тих біомолекул, які необхідні для виконання специфічних функцій клітини та організму.
Метаболічний шлях – це послідовність хімічних перетворень конкретної речовини у організмі. Проміжні продукти, що утворюються в процесі перетворення, називають метаболітами, а остання сполука метаболічного шляху – кінцевим продуктом. Прикладом метаболічного шляху є гліколіз, синтез холестерину.
Метаболічний цикл – це такий метаболічний шлях, один із кінцевих продуктів якого ідентичний одному із сполук залучених до цього процесу. Найбільш важливими в організмі людини метаболічними циклами є цикл трикарбонових кислот (цикл Кребса) та орнітиновий цикл сечовиноутворення.
Майже всі метаболічні реакції зрештою пов'язані між собою, оскільки продукт однієї ферментативної реакції служить субстратом для іншої, яка в цьому процесі відіграє роль наступної стадії. Таким чином, метаболізм можна подати у вигляді надзвичайно складної мережі ферментативних реакцій. Якщо потік поживних речовин в одній частині цієї мережі зменшиться або порушиться, то у відповідь можуть відбутися зміни в іншій частині мережі, для того щоб ця перша зміна була якось врівноважена або компенсована. Більш того, і катаболічні та анаболічні реакції відрегульовані таким чином, щоб вони протікали найбільше економічно, тобто з найменшою витратою енергії та речовин. Наприклад, окислення поживних речовин у клітині відбувається зі швидкістю, якраз достатньою для того, щоб задовольнити її енергетичні потреби в даний момент.
Специфічні та загальні шляхи катаболізму
У катаболізмі розрізняють три стадії:
1. Полімери перетворюються на мономери (білки – на амінокислоти, вуглеводи на моносахариди, ліпіди – на гліцерол та жирні кислоти). Хімічна енергія у своїй розсіюється як тепла.
2. Мономери перетворюються на загальні продукти, у переважній більшості на ацетил-КоА. Хімічна енергія частково розсіюється як тепла, частково накопичується як відновлених коферментних форм (НАДН, ФАДН2), частково запасається у макроергічних зв'язках АТФ (субстратное фосфорилирование).
Перша і друга стадії катаболізму відносяться до специфічних шляхів, які унікальні для метаболізму білків, ліпідів і вуглеводів.
3. Заключний етап катаболізму, зводиться до окислення ацетил-КоА до СО 2 і Н 2 Про реакціях циклу трикарбонових кислот (циклу Кребса) – загальний шлях катаболізму. Окисні реакції загального шляху катаболізму пов'язані з ланцюгом тканинного дихання. При цьому енергія (40-45%) запасається у вигляді АТФ (окисне фосфорилювання).
В результаті специфічних та загальних шляхів катаболізму біополімери (білки, вуглеводи, ліпіди) розпадаються до СО 2 , Н 2 Про і NH 3 які є основними кінцевими продуктами катаболізму.
Метаболіти в нормі та при патології
У живій клітині щомиті утворюються сотні метаболітів. Однак їх концентрації підтримуються на певному рівні, що є специфічною біохімічною константою або референтною величиною. За хвороб відбувається зміна концентрації метаболітів, що є основою біохімічної лабораторної діагностики. До нормальних метаболітів відносять глюкозу, сечовину, холестерол, загальний білок сироватки крові та низку інших. Вихід концентрації цих речовин межі фізіологічних норм (підвищення чи зниження) говорить про порушення їх обміну в організмі. Більше того, ряд речовин в організмі здорової людини виявляється лише у певних біологічних рідинах, що обумовлюється специфікою їхнього метаболізму. Наприклад, білки сироватки крові в нормі не проходять через нирковий фільтр і, відповідно, не виявляються у сечі. Але при запаленні нирок (гломерулонефриті) білки (насамперед альбуміни) проникають через капсулу клубочка, з'являються у сечі – протеїнурія та трактуються як патологічні компоненти сечі.
Патологічними метаболітами є мієломні білки (білки Бенс-Джонса), парапротеїни при макроглобулінемії Вальденштрема, накопичення аномального глікогену при глікогенозах, різноманітних складних фракцій ліпідів при сфінголіпідозах і т.д. Вони виявляються лише за хвороб і для здорового організму не характерні.
Рівні вивчення обміну речовин
Рівні вивчення обміну речовин:
1. Цілий організм.
2. Ізольовані органи (перфузовані).
3. Зрізи тканин.
4. Культури клітин.
5. Гомогенати тканин.
6. Ізольовані клітинні органели.
7. Молекулярний рівень (очищені ферменти, рецептори тощо).
Досить часто вивчення метаболізму використовують радіоактивні ізотопи (3 H, 32 P, 14 C, 35 S, 18 O), якими помічають речовини, що вводяться в організм. Потім можна простежити клітинну локалізацію цих речовин, визначити період напіврозпаду та їх метаболічні шляхи.
Мал. 8.1. Схема специфічних та загальних шляхів катаболізму
Розділ 9. Біологічні мембрани
Клітина представляє біологічну систему, основу якої складають мембранні структури, що відокремлюють клітину від зовнішнього середовища, що формують її відсіки (компартменти), а також забезпечують надходження та видалення метаболітів, сприйняття та передачу сигналів і є структурними організаторами метаболічних шляхів.
Узгоджене функціонування мембранних систем – рецепторів, ферментів, транспортних механізмів допомагає підтримувати гомеостаз клітини та водночас швидко реагувати зміни зовнішнього середовища.
Мембрани – нековалентні надмолекулярні структури. Білки та ліпіди у них утримуються разом безліччю нековалентних взаємодій (кооперативних за характером).
До основних функцій мембран можна віднести:
1. відділення клітини від довкілля та формування внутрішньоклітинних компартментів (відсіків);
2. контроль та регулювання транспорту великого розмаїття речовин через мембрани (виборча проникність);
3. участь у забезпеченні міжклітинних взаємодій;
4. сприйняття та передача сигналу всередину клітини (рецепція);
5. локалізація ферментів;
6. Енерготрансформуюча функція.
Мембрани асиметричні у структурному та функціональному відносинах (вуглеводи локалізуються завжди зовні і їх немає на внутрішній стороні мембрани). Це динамічні структури: білки і ліпіди, що входять до їх складу, можуть рухатися в площині мембрани (латеральна дифузія). Однак існує і перехід білків і ліпідів з одного боку мембрани на іншу (поперечна дифузія, фліп-флоп), яка відбувається вкрай повільно. Рухливість і плинність мембран залежить від її складу: співвідношенням насичених і ненасичених жирних кислот, і навіть холестеролу. Плинність мембрани тим нижча, чим вище насиченість жирних кислот у фосфоліпідах і чим більший вміст холестеролу. Крім того, для мембран характерна самоскладання.
Загальні властивості клітинних мембран:
1. легко проникні для води та нейтральних ліпофільних сполук;
2. меншою мірою проникні для полярних речовин (цукри, аміди);
3. погано проникні для невеликих іонів (Na +, Cl - та ін);
4. характерно високий електричний опір;
5. асиметричність;
6. можуть спонтанно відновлювати цілісність;
7. рідинність.
Хімічний склад мембран.
Мембрани складаються з ліпідних та білкових молекул, відносна кількість яких у різних мембран широко коливається. Вуглеводи містяться у формі глікопротеїнів, гліколіпідів та становлять 0,5%-10% речовин мембрани. Відповідно до рідинно-мозаїчної моделі будови мембрани (Сенджер і Ніколсон, 1972 р.) основу мембрани становить подвійний ліпідний шар, у формуванні якого беруть участь фосфоліпіди та гліколіпіди. Ліпідний бислой утворений двома рядами ліпідів, гідрофобні радикали яких заховані всередину, а гідрофільні групи звернені назовні та контактують з водним середовищем. Білкові молекули ніби розчинені в ліпідному бісла і відносно вільно «плавають у ліпідному морі у вигляді айсбергів, на яких ростуть дерева глікоколіксу».
Ліпіди мембран.
Мембранні ліпіди – амфіфільні молекули, тобто. в молекулі є як гідрофільні групи (полярні головки), так і аліфатичні радикали (гідрофобні хвости), які мимовільно формують бислой, в якому хвости ліпідів звернені один до одного. Товщина одного ліпідного шару 2,5 нм, у тому числі 1 нм посідає головку і 1,5 нм на хвіст. У мембранах присутні три основні типи ліпідів: фосфоліпіди, гліколіпіди та холестерол. Середнє молярне відношення холестерол/фосфоліпіди дорівнює 0,3-0,4, але в плазматичній мембрані це співвідношення набагато вище (0,8-0,9). Наявність холестеролу в мембранах зменшує рухливість жирних кислот, знижує латеральну дифузію ліпідів та білків.
Фосфоліпіди можна розділити на гліцерофосфоліпіди та сфінгофосфоліпіди. Найбільш поширені гліцерофосфоліпіди мембран – фосфатидилхоліни та фосфатидилетаноламіни. Кожен гліцерофосфоліпід, наприклад фосфатидилхолін, представлений декількома десятками фосфатидилхолінів, що відрізняються один від одного будовою жирнокислотних залишків.
Перед гліцерофосфоліпідів припадає 2–8% всіх фосфоліпідів мембран. Найбільш поширеними є фосфатидилінозитоли.
Специфічні фосфоліпіди внутрішньої мембрани мітохондрій – кардіоліпіни (дифосфатидглицероли), побудовані на основі гліцеролу та двох залишків фосфатидної кислоти, становлять близько 22% від усіх фосфоліпідів мітохондріальних мембран.
У мієліновій оболонці нервових клітин у значних кількостях містяться сфінгомієліни.
Гліколіпіди мембран представлені цереброзидами та гангліозидами, в яких гідрофобна частина представлена церамідом. Гідрофільна група – вуглеводний залишок – глікозидним зв'язком приєднаний до гідроксильної групи першого вуглецевого атома цераміду. У значних кількостях гліколіпіди знаходяться у мебранах клітин мозку, епітелію та еритроцитів. Гангліозиди еритроцитів різних індивідуумів відрізняються будовою олігосахаридних ланцюгів та виявляють антигенні властивості.
Холестерол присутній у всіх мембранах тваринних клітин. Його молекула складається з жорсткого гідрофобного ядра та гнучкого вуглеводневого ланцюга, єдина гідроксильна група є полярною головкою.
Функції мембранних ліпідів.
Фосфо- і гліколіпіди мембран, крім участі у формуванні ліпідного бислоя, виконують низку інших функцій. Ліпіди мембран формують середовище для функціонування мембранних білків, які приймають у ній нативну конформацію.
Деякі мембранні ліпіди – попередники вторинних посередників під час передачі гормональних сигналів. Так фосфатидилінозитолдифосфат під дією фосфоліпази С гідролізується до діацилгліцеролу та інозитолтрифосфату, які є вторинними посередниками гормонів.
Ряд ліпідів бере участь у фіксації заякорених білків. Прикладом заякореного білка є ацетилхолінестераза, яка фіксується на постсинаптичній мембрані до фосфатитилінозитолу.
Білки мембран.
Мембранні білки відповідають за функціональну активність мембран і їх частку припадає від 30 до 70%. Білки мембран відрізняються за своїм становищем у мембрані. Вони можуть глибоко проникати в ліпідний бішар або навіть пронизувати його - інтегральні білки, різними способами прикріплюватися до мембрани - поверхневі білки, або ковалентно контактувати з нею - заякорені білки. Поверхневі білки майже завжди глікозильовані. Олігосахаридні залишки захищають білок від протеолізу, беруть участь у впізнанні лігандів та адгезії.
Білки, локалізовані в мембрані, виконують структурну та специфічні функції:
1. транспортну;
2. ферментативну;
3. рецепторну;
4. антигенну.
Механізми мембранного транспорту речовин
Розрізняють кілька способів перенесення речовин через мембрану:
1. Проста дифузія– це перенесення невеликих нейтральних молекул за градієнтом концентрації без витрат енергії та переносників. Найлегше проходять простою дифузією через ліпідну мембрану малі неполярні молекули, такі як О2, стероїди, тиреоїдні гормони. Малі полярні незаряджені молекули – СО 2 , NH 3 , H 2 O, етанол та сечовина – також дифундують із достатньою швидкістю. Дифузія гліцеролу йде значно повільніше, а глюкоза практично не здатна самостійно пройти через мембрану. Для всіх заряджених молекул незалежно від розміру ліпідна мембрана не проникна.
2. Полегшена дифузія- Перенесення речовини за градієнтом концентрації без витрат енергії, але з переносником. Характерна для водорозчинних речовин. Полегшена дифузія відрізняється від простої більшої швидкості перенесення і здатність до насичення. Розрізняють два різновиди полегшеної дифузії:
Транспорт спеціальними каналами, утвореними в трансмебранних білках (наприклад, катіонселективні канали);
За допомогою білків-транслоказів, які взаємодіють зі специфічним лігандом, забезпечують його дифузію за градієнтом концентрації (пінг-понг) (перенесення глюкози в еритроцити за допомогою білка-переносника ГЛЮТ-1).
Кінетично перенесення речовин полегшеною дифузією нагадує ферментативну реакцію. Для транслоказ існує концентрація, що насичує ліганда, при якій всі центри зв'язування білка з лігандом зайняті, і білки працюють з максимальною швидкістю. Тому швидкість транспорту речовин полегшеною дифузією залежить не тільки від градієнта концентрацій речовини, що переноситься, але і від кількості беків-переносників у мембрані.
Проста та полегшена дифузія відноситься до пасивного транспорту, оскільки відбувається без витрати енергії.
3. Активний транспорт– транспорт речовини проти градієнта концентрації (незаряджені частинки) або електрохімічного градієнта (для заряджених частинок), що потребує витрат енергії, найчастіше АТФ. Виділяють два види його: первинно активний транспорт використовує енергію АТФ або окислювально-відновного потенціалу та здійснюється за допомогою транспортних АТФ-аз. Найбільш поширені в плазматичній мембрані клітин людини Na+, K+-АТФ-аза, Са2+-АТФ-аза, Н+-АТФ-аза.
При вторинно активному транспорті використовується градієнт іонів, створений на мембрані за рахунок роботи системи первинно активного транспорту (всмоктування глюкози клітинами кишечника та реабсорбція з первинної сечі глюкози та амінокислот клітинами нирок, що здійснюються при русі іонів Na + за градієнтом концентрації).
Перенесення через мембрану макромолекул. Транспортні білки забезпечують перенесення через клітинну мембрану полярних молекул невеликого розміру, але вони не можуть транспортувати макромолекули, наприклад, білки, нуклеїнові кислоти, полісахариди або окремі частинки.
Механізми, за допомогою яких клітини можуть засвоювати такі речовини або видаляти їх із клітини, відрізняються від механізмів транспортування іонів та полярних сполук.
1. Ендоцитоз.Це перенесення речовини із середовища в клітину разом із частиною плазматичної мембрани. Шляхом ендоцитозу (фагоцитозу) клітини можуть поглинати великі частки, такі як віруси, бактерії чи фрагменти клітин. Поглинання рідини та розчинених у ній речовин за допомогою невеликих бульбашок називають піноцитозом.
2. Екзоцитоз. Макромолекули, наприклад, білки плазми, пептидні гормони, травні ферменти синтезуються в клітинах і потім секретуються в міжклітинний простір або кров. Але мембрана не проникна таких макромолекул чи комплексів, їх секреція відбувається шляхом екзоцитозу. В організмі є як регульований так і не регульований шлях екзоцитозу. Нерегульована секреція характеризується безперервним синтезом білків, що секретуються. Прикладом може бути синтез та секреція колагену фібробластами для формування міжклітинного матриксу.
Для регульованої секреції характерні зберігання приготовлених експорту молекул в транспортних бульбашках. За допомогою регульованої секреції відбуваються виділення травних ферментів, а також секреція гормонів та нейромедіаторів.
Розділ 10. Енергетичний обмін. Біологічне окиснення
Живі організми з погляду термодинаміки – відкриті системи. Між системою та довкіллям можливий обмін енергії, який відбувається відповідно до законів термодинаміки. Кожна органічна сполука, що надходить в організм, має певний запас енергії (Е). Частина цієї енергії може бути використана для здійснення корисної роботи. Таку енергію називають вільною енергією (G). Напрямок хімічної реакції визначається значенням DG. Якщо ця величина негативна, реакція протікає мимовільно. Такі реакції називаються екзергонічними. Якщо DG позитивно, то реакція протікатиме лише при надходженні вільної енергії ззовні – це ендергонічні реакції. У біологічних системах термодинамічно невигідні ендергонічні реакції можуть протікати лише за рахунок енергії екзергонічних реакцій. Такі реакції називають енергетично сполученими.
Найважливішою функцією багатьох біологічних мембран служить перетворення однієї форми енергії на іншу. Мембрани, що мають такі функції, називаються енергоперетворювальними. Будь-яка мембрана, що виконує енергетичну функцію, здатна до перетворення хімічної енергії субстратів, що окислюються, або АТФ в електричну енергію, а саме в трансмембранну різницю електричних потенціалів (DY) або в енергію різниці концентрацій речовин, що містяться в розділених мембраною розчинах, і навпаки. Серед енергоперетворювальних мембран, що мають найбільше значення, можна назвати внутрішню мембрану мітохондрій, зовнішню цитоплазматичну мембрану, мембрани лізосом та комплексу Гольджі, саркоплазматичний ретикулум. Зовнішня мембрана мітохондрій та ядерна мембрана не може перетворювати одну форму енергії на іншу.
Перетворення енергії в живій клітині описується такою загальною схемою:
Енергетичні ресурси → ΔμI → робота
де ? Тому даний іон може бути названий іоном, що сполучає. Основним сполучним іоном в клітині еукаріотів є Н + , і відповідно ΔμН + є основною конвертованою формою запасання енергії. Другим за значимістю іоном, що сполучає, є Na + (ΔμNa +). У той час як Сa 2+ , K + і Cl - не використовуються для будь-якої роботи.
Біологічне окислення – це процес дегідрування субстрату за допомогою проміжних переносників водню та його кінцевого акцептора. Якщо ролі кінцевого акцептора виступає кисень, процес називається аеробним окисленням чи тканинним диханням, якщо кінцевий акцептор представлений не киснем – анаеробним окисленням. Анаеробне окислення має обмежене значення в організмі людини. Основна функція біологічного окислення – забезпечення клітини енергією у доступній формі.
Тканинне дихання – процес окислення водню киснем до води ферментами ланцюга тканинного дихання. Воно протікає за такою схемою:

Речовина окислюється, якщо віддає електрони або одночасно електрони та протони (атоми водню), або приєднує кисень. Здатність молекули віддавати електрони іншій молекулі визначається окислювально-відновним потенціалом (редокс-потенціалом). Будь-яке з'єднання може віддавати електрони тільки речовині з вищим окислювально-відновним потенціалом. Окислювач і відновник завжди утворюють пару.
Виділяють 2 типи окислюваних субстратів:
1. Піридінзалежні – спиртові або альдегідні – ізоцитрат, α-кетоглутарат, піруват, малат, глутамат, β-гідроксіацил-КоА, β-гідроксибутират, – в їх дегідруванні беруть участь НАД-залежні дегідрогенази.
2. флавінзалежні - є похідними вуглеводнів - сукцинат, ацил-КоА, гліцерол-3-фосфат, холін - при дегідруванні передають водень на ФАД-залежні дегідрогенази.
Ланцюг тканинного дихання – послідовність переносників протонів водню (Н+) та електронів від окислюваного субстрату на кисень, локалізованих на внутрішній мембрані мітохондрій.

Мал. 10.1. Схема ЦТД
Компоненти ЦТД:
1. НАД-залежні дегідрогенази дегідрують піридинзалежні субстрати і акцептують 2 і один Н + .
2. ФАД (ФМН) - залежні дегідрогенази акцептують 2 атоми водню (2Н + і 2?). ФМН – залежна дегідрогеназа дегідрує тільки НАДН, тоді як ФАД-дегідрогенази окислюють флавінзалежні субстрати.
3. Жиророзчинний переносник убихинон (кофермент Q, КоQ) – вільно переміщається мембраною мітохондрій і акцептує два атоми водню і перетворюється на КоQH 2 (відновлена форма – убихинол).
4. Система цитохромів – переносить лише електрони. Цитохроми залізовмісні білки, простетична група яких структурою нагадує гем. На відміну від гему, атом заліза в цитохромі може оборотно переходити з двох – у тривалентний стан (Fe 3+ + ē → Fe 2+). Це забезпечує участь цитохрому в транспорті електронів. Цитохроми діють у порядку зростання їх редокс-потенціалу і в дихальному ланцюгу розташовуються наступним чином: b-с 1 -с-а-а 3 . Два останні працюють в асоціації як один фермент цитохромоксидазу аа3. Цитохромоксидаза складається з 6 субодиниць (2 - цитохрому а і 4 - цитохрому а 3). У цитохромі а 3 крім заліза є атоми міді, і він передає електрони безпосередньо на кисень. Атом кисню при цьому заряджається негативно і набуває здатності взаємодіяти з протонами з утворенням метаболічної води.
Залізосерні білки (FeS) – містять негемове залізо та беруть участь в окислювально-відновних процесах, що протікають за одноелектронним механізмом та асоційовані з флавопротеїнами та цитохромом b.
Структурна організація ланцюга тканинного дихання
Компоненти дихального ланцюга у внутрішній мембрані михохондрій формують комплекси:
1. I комплекс (НАДН-КоQН2-редуктаза) – приймає електорони від мітохондріального НАДН та транспортує їх на КоQ. Протони транспортуються у міжмембранний простір. Проміжним акцептором та переносником протонів та електронів є ФМН та залізосерні білки. I комплекс поділяє потік електронів та протонів.
2. II комплекс – сукцинат – КоQ – редуктаза – включає ФАД-залежні дегідрогенази та залізосерні білки. Він транспортує електрони та протони від флавінзалежних субстратів на убихинон, з утворенням проміжного ФАДН 2 .
Убіхінон легко переміщається мембраною і передає електрони на III комплекс.
3. III комплекс - КоQН 2 - цитохром с - редуктаза - має у своєму складі цитохроми b і с 1, а також залізосерні білки. Функціонування КоQ з III комплексом призводить до поділу потоку протонів і електронів: протони з матриксу перекачуються в міжмембранний простір мітохондрій, а електрони транспортуються далі ЦТД.
4. IV комплекс – цитохром а - цитохромоксидаза – містить цитохромоксидазу і транспортує електрони на кисень з проміжного переносника цитохрому, який є рухомим компонентом ланцюга.
Існує 2 різновиди ЦТД:
1. Повний ланцюг - до нього вступають пиридинзависимые субстрати і здають атоми водню на НАД-залежні дегідрогенази
2. Неповна (укорочена або редукована) ЦТД в якій атоми водню передаються від ФАД-залежних субстратів, в обхід першого комплексу.
Окисне фосфорилювання АТФ
Окисне фосфорилювання - процес утворення АТФ, пов'язаний з транспортом електронів по ланцюгу тканинного дихання від субстрату, що окислюється, на кисень. Електрони завжди прагнуть переходити від електронегативних систем до електропозитивних, тому їх транспорт ЦТД супроводжується зниженням вільної енергії. У дихальному ланцюгу кожному етапі зниження вільної енергії відбувається ступінчасто. При цьому можна виділити три ділянки, в яких перенесення електронів супроводжується великим зниженням вільної енергії. Ці етапи здатні забезпечити енергією синтез АТФ, оскільки кількість вільної енергії, що виділяється, приблизно дорівнює енергії, необхідної для синтезу АТФ з АДФ і фосфату.
Для пояснення механізмів сполучення дихання та фосфорилювання висунутий ряд гіпотез.
Механохімічна чи конформаційна (Грін-Бойєра).
У процесі перенесення протонів та електронів змінюється конформація білків-ферментів. Вони переходять у новий, багатий на енергію конформаційний стан, а потім при поверненні у вихідну конформацію віддають енергію для синтезу АТФ.
Гіпотеза хімічної пари (Ліпмана).
У поєднанні дихання та фосфорилювання беруть участь «сполучні» речовини. Вони акцептують протони та електрони та взаємодіють з Н 3 РО 4 . У момент віддачі протонів та електронів зв'язок з фосфатом стає макроергічною та фосфатна група передається на АДФ з утворенням АТФ шляхом субстратного фосфорилювання. Гіпотеза логічна, проте досі не виділені «сполучні» речовини.
Хеміоосмотична гіпотеза Пітера Мітчелла (1961р.)
Основні постулати цієї теорії:
1. внутрішня мембрана мітохондрій непроникна для іонів Н + і ОН −;
2. за рахунок енергії транспорту електронів через I, III та IV комплекси дихального ланцюга з матриксу викачуються протони;
3. електрохімічний потенціал, що виникає на мембрані, є проміжною формою запасання енергії;
4. повернення протонів в матрикс мітохондрії через протонний канал АТФ синтази є постачальником енергії для синтезу АТФ за схемою
АДФ+Н 3 РВ 4 →АТФ+Н 2 О
Докази хеміоосмотичної теорії:
1. на внутрішній мембрані є градієнт Н + і його можна виміряти;
2. створення градієнта Н+ у мітохондрії супроводжується синтезом АТФ;
3. іонофори (роз'єднувачі), що руйнують протонний градієнт, гальмують синтез АТФ;
4. інгібітори, що блокують транспорт протонів протонними каналами АТФ-синтази, інгібують синтез АТФ.
Будова АТФ-синтази
АТФ-синтаза – інтегральний білок внутрішньої мембрани мітохондрій. Він розташований у безпосередній близькості до дихального ланцюга та позначається як V комплекс. АТФ-синтазу складається з 2 субодиниць, що позначаються як F 0 і F 1 . Гідрофобний комплекс F 0 занурений у внутрішню мембрану мітохондрій і складається з декількох протомерів, що утворюють канал, по якому протони переносяться в матрикс. Субодина F 1 виступає в мітохондріальний матрикс і складається з 9 протомерів. Причому три з них пов'язують субодиниці F 0 і F 1 утворюючи своєрідну ніжку і є чутливими до олігоміцину.
Суть хеміоосмотичної теорії: за рахунок енергії перенесення електронів по ЦТД відбувається рух протонів через внутрішню мітохондріальну мембрану в міжмембранний простір, де створюється електрохімічний потенціал (ΔμН+), який призводить до конформаційної перебудови активного центру АТФ-синтази, в результаті чого стає можливим зворотний через протонні канали АТФ-синтази. При поверненні протонів назад електрохімічний потенціал трансформується на енергію макроергічного зв'язку АТФ. АТФ, що утворилася, за допомогою білка-переносника транслокази переміщається в цитозоль клітини, а замість матрикс надходять АДФ і Ф н.
Коефіцієнт фосфорилювання (Р/О) – число атомів неорганічного фосфату, включених у молекули АТФ, у перерахунку однією атом використаного поглиненого кисню.
Пункти фосфорилювання – ділянки у дихальному ланцюгу, де енергія транспорту електоронів використовується на генерацію протонного градієнта, а потім у ході фосфорилювання запасається у формі АТФ:
1. 1 пункт – між піридинзалежною та флавінзалежною дегідрогеназами; 2 пункт - між цитохромами b і з 1; 3 пункт – між цитохромами а та а 3 .
2. Отже, при окисленні НАД-залежних субстратів коефіцієнт Р/О дорівнює 3, оскільки електрони від НАДН транспортуються за участю всіх комплексів ЦТД. Окислення ФАД-залежних субстратів йде в обхід I комплексу дихального ланцюга та Р/О дорівнює 2.
Порушення енергетичного обміну
Усі живі клітини постійно потребують АТФ реалізації різних видів діяльності. Порушення будь-якого етапу метаболізму, що призводять до припинення синтезу АТФ, є згубними для клітини. Тканини з високими енергетичними потребами (ЦНС, міокард, нирки, скелетні м'язи та печінка) є найбільш вразливими. Стани, за яких синтез АТФ знижений поєднують терміном «гіпоенергетичні». Причини цих станів можна розбити на дві групи:
Аліментарні – голодування та гіповітамінози В2 та РР – виникає порушення постачання окислюваних субстратів у ЦТД або синтез коферментів.
Гіпоксичні – виникають у разі порушення доставки чи утилізації кисню у клітині.
Регулювання ЦТД.
Здійснюється з допомогою дихального контролю.
Дихальний контроль – це регуляція швидкості перенесення електронів по дихальному ланцюзі ставленням АТФ/АДФ. Чим менше це відношення, тим інтенсивніше йде дихання та активніше синтезується АТФ. Якщо АТФ немає, та її концентрація у клітині зростає, то припиняється потік електронів до кисню. Накопичення АДФ збільшує окислення субстратів та поглинання кисню. Механізм дихального контролю характеризується високою точністю і має важливе значення, тому що в результаті його дії швидкість синтезу АТФ відповідає потребам клітини енергії. Запасів АТФ у клітині немає. Відносні концентрації АТФ/АДФ у тканинах змінюються у вузьких межах, тоді як споживання енергії клітиною може змінюватись у десятки разів.
Американський біохімік Д. Чанс запропонував розглядати 5 станів мітохондрій, при яких швидкість їхнього дихання обмежується певними факторами:
1. Нестача SH 2 та АДФ – швидкість дихання дуже низька.
2. Нестача SH 2 за наявності АДФ – швидкість обмежена.
3. Є SH 2 та АДФ – дихання дуже активно (лімітується лише швидкістю транспорту іонів через мембрану).
4. Нестача АДФ за наявності SH 2 - дихання гальмується (стан дихального контролю).
5. Нестача кисню, за наявності SH 2 та АДФ – стан анаеробіозу.
Мітохондрії в клітці знаходяться в стані 4, при якому швидкість дихання визначається кількістю АДФ. Під час посиленої роботи можуть перебувати у стані 3 (вичерпуються можливості дихального ланцюга) або 5 (недолік кисню) – гіпоксії.
Інгібітори ЦТД – це лікарські препарати, які блокують перенесення електронів ЦТД. До них відносять: барбітурати (амітал), що блокують транспорт електронів через I комплекс дихального ланцюга, антибіотик антиміцин блокує окислення цитохрому b; монооксид вуглецю та ціаніди інгібують цитохромооксидазу та блокують транспорт електронів на кисень.
Інгібітори окисного фосфорилювання (олігоміцин) – це речовини, які блокують транспорт Н+ протонним каналом АТФ-синтази.
Роз'єднувачі окисного фосфорилювання (іонофори) - це речовини, які пригнічують окисне фосфорилювання, не впливаючи при цьому на процес перенесення електронів ЦТД. Механізм дії роз'єднувачів зводиться до того, що вони є жиророзчинними (ліпофільними) речовинами і мають здатність зв'язувати протони і переносити їх через внутрішню мембрану мітохондрій в матрикс, минаючи протонний канал АТФ-синтази. Енергія, що виділяється при цьому, розсіюється у вигляді тепла.
Штучні роз'єднувачі – динітрофенол, похідні вітаміну К (дикумарол), деякі антибіотики (валіноміцин).
Природні роз'єднувачі – продукти перекисного окислення ліпідів, жирні кислоти з довгим ланцюгом, великі дози йодовмісних гормонів щитовидної залози, білки термогеніни.
На роз'єднанні дихання та фосфорилювання базується терморегуляторна функція тканинного дихання. Мітохондрії бурої жирової тканини продукують більше тепла, тому що присутній у них білок термогенін роз'єднує окислення та фосфорилювання. Це має важливе значення у підтримці температури тіла новонароджених.
Біохімічні шляхи адаптації тварин і людини до геохімічних умов середовища - це питання не про окремі ізольовані організми, а про пристосовність особин усередині популяції як її частини. Не тільки популяцію, а й особину необхідно розглядати як фізіолого-генетичні одиниці еволюційного процесу, що займають різні рівні у структурі біосфери. Вивчення внутрішньопопуляційної мінливості організмів розкриває роль популяції в еволюції окремих особин і своєю чергою окремих особин в еволюції популяції як цілого.
Значення природного геохімічного середовища для розвитку організмів та еволюції життя визначається використанням організмами багатьох хімічних елементів у процесах обміну речовин та біологічно активних сполук, до складу яких входять ці елементи. Тому неоднорідність геохімічного середовища є однією з важливих причин мінливості обміну речовин та синтезу біологічно активних сполук в організмах.
Сучасними біогеохімією, геохімією, ґрунтознавством з повною очевидністю доведено геохімічну неоднорідність біосфери, значну хімічну мінливість середовища життя в умовах літосфери, гідросфери та тропосфери. Організми глибоко пов'язані з геохімічним середовищем. Вони поглинають із цього середовища всі доступні хімічні елементи, що утворюють розчинні сполуки, або активно перетворюють нерозчинні сполуки на доступні.
Дуже важливою властивістю біосфери є єдність геохімічного середовища та життя, що склалося в процесі еволюції біосфери та виражається постійною залежністю життя від геохімічних умов середовища та клімату (водний режим, температура, інсоляція, конвекція). Ступінь накопичення хімічних елементів організмами визначається не тільки геохімією середовища та біологічною природою організмів, а й біогеохімічними харчовими ланцюгами, що складаються із системи взаємозалежних ланок, через які здійснюється зв'язок організмів та середовища (грунтоутворюючі породи, ґрунти, води, повітря, мікроорганізми, рослини, тварини, людина).
Для з'ясування складних шляхів пристосування організмів до геохімічного середовища необхідно визначити загальні закономірності на організм різних концентрацій хімічних елементів. Встановлено, що як за браку, і надлишку в раціоні кобальту, міді, цинку приріст ваги тварин зменшується. Синтез багатьох біологічно активних сполук в організмі також залежить від вмісту певних мікроелементів у раціоні чи геохімічному середовищі. Накопичення вітаміну В 12 у печінці тварин (вівці, велика рогата худоба, кролики та інші) значно знижується при зменшенні кількості кобальту в середовищі або раціоні та незначно високому його вмісті. Нестача йоду або його надлишок призводять до компенсаторного збільшення розмірів щитовидної залози і гальмування синтезу в ній сполук, що містять йод: йодтирозинів, йодтиронінів, тироксину та їх похідних. Важливими регуляторами пуринового обміну є молібден та мідь. При відносному нестачі або надлишку молібдену у людини і тварин гальмується синтез або зменшується активність ферменту ксантиноксидази, що містить молібден, що перетворює деякі пуринові основи на сечову кислоту, а у тварин при низькій, як і при підвищеній кількості міді знижується синтез уратоксидази, що окислює сечову кислоту в . Можна навести багато таких прикладів. У всіх випадках спостерігається та сама закономірність: недолік чи надлишок серед чи раціоні тих чи інших хімічних елементів гальмує певні біохімічні процеси в організмі. Отже, рівень синтезу ферментів та інших біологічно активних сполук в організмі, що забезпечує нормальний перебіг життєвих процесів, спостерігається лише при певних концентраціях та співвідношеннях у середовищі та організмі мікроелементів (рисунок). Регулюючі системи організму (депонування, видільна, бар'єрна, синтезу біологічно активних сполук та інші) не можуть нормально функціонувати при концентрації мікроелементів вище або нижче цих меж.
У таких випадках відбувається порушення функцій або зрив їх, виникають дисфункції, аномалії розвитку та обміну речовин, що може призвести до появи ендемічних захворювань у людини і тварин.
Окремі реакції на нестачу чи надлишок мікроелементів ми розглядаємо як частину цілого. Для розуміння цих явищ необхідно з'ясувати основні точки докладання хімічних елементів до біохімічних процесів і встановити ланцюг процесів, що залучають цілий організм до реакції на недолік або надлишок певних елементів, тобто пояснити значення частини реакції цілого. Наприклад, основна роль кобальту в організмі полягає в мікробному синтезі в травному тракті вітаміну В 12 містить кобальт.
При нестачі кобальту може значно послаблюватися синтез вітаміну B 1 2 та його всмоктування через слизову оболонку кишечника. Ослаблення процесу всмоктування вітаміну B 12 обумовлюється зниженням секреції шлункового соку і недоліком мукопротеїду, що дає з вітаміном В 12 з'єднання, для якого проникна слизова оболонка кишечника. Ще немає ясності в питанні про значення для усмоктування вітаміну B 12 утворення його сполуки з цинком. Внаслідок нестачі в організмі вітаміну В 12 зменшується його депонування печінкою та іншими органами. Це особливо яскраво виражено у жуйних тварин (вівці, велика рогата худоба), у яких недолік кобальту викликає захворювання ендемічними гіпо-і авітамінозом В 12, але досить ясно проявляється н в інших групах (кролики, свині). За нестачі в організмі вітаміну В 12 розвивається перніціозна анемія. Крім того, при цьому гальмується синтез коензиму ізомерази, що діє на метилмалонову кислоту, і порушується метилювання тиміну та метіоніну. Порушення таких важливих процесів в організмі призводить до включення в патологічний процес нових і нових ланок обміну речовин. Недостатність кобальту викликає послаблення синтезу білків, нуклеїнових кислот (наприклад, тимонуклеїнової кислоти), зниження активності багатьох ферментів (наприклад, аргінази), зокрема ферментів нуклеїнового обміну (ДНК-ази), зниження основного обміну, втрату ваги тварин. Ще недостатньо вивчено значення кобальту у процесах обміну речовин. Але безперечно, що недостатність кобальту може спричинити глибокі зміни цілого організму.
При малій концентрації міді у навколишньому середовищі та раціоні у тварин спостерігаються характерні зміни обміну речовин, що викликаються зниженням активності окисних ферментів, що містять мідь або залізо. Це особливо чітко проявляється в біогеохімічних провінціях ендемічної атаксії, в яких нестача міді поєднується з надлишком молібдену та сульфатів. При нестачі міді пригнічення цитохромоксидази у щурів сильніше виражене в печінці, у свиней та курчат – у серці, у овець – у мозку, а пригнічені сукцинатдегідрогенази у овець – у білій речовині головного та спинного мозку; гальмування активності НАД-цитохром-с-редуктази встановлено в мітохондріях печінки щурів, сульфідоксидази - в печінці овець, ДОФА-оксидази - в печінці та сірій речовині головного та спинного мозку овець, моноаміноксидази у свиней та овець - у сироватці крові, у аорті; активність ізоцитратдегідрогенази мітохондрій печінки щура при нестачі міді підвищується, оскільки, мабуть, мідь легко пригнічує цей фермент; при нестачі міді та підвищеному вмісті молібдену посилюється активність ферментів пуринового обміну - ксантиноксидази та уратоксидази у різних видів ссавців.
Наведені дані про вплив нестачі міді на активність багатьох ферментів різних органів та тканин показують значення порівняльних фізіологічних досліджень.
Зміна активності ферментів тваринного організму при нестачі міді спричиняє порушення багатьох біохімічних процесів та фізіологічних станів. Цілком ймовірно, що важливими токсичними агентами в організмі при нестачі міді є сульфіди, що утворюються в печінці при розпаді пістеїну і в жуйних рубці в результаті мікробіологічного відновлення сульфатів. При надлишку в раціоні сульфатів утворюються сульфіди, при цьому харчова мідь перетворюється на сірчисту, не засвоювану організмом, що посилює дефіцит міді. Знешкодження великої частини сульфідів відбувається в печінці шляхом їх окислення в тіосульфат та сульфат. Окислення сульфідів в тіосульфат здійснюється сульфідоксидазою печінки, активність якої пригнічується при низькому вмісті міді в раціоні та надлишку молібдену. Очевидно, при недостатності міді посилюється утворення сульфідів у тканинах, а надлишок сульфідів ще більше посилює її. Досліди з радіоактивною міддю дозволяють вважати, що в біогеохімічних провінціях при нестачі міді та надлишку молібдену та сульфатів збільшується втрата міді тканинами, що також посилює дефіцит цього мікроелемента.
Нестача міді, змінюючи активність багатьох ферментів, викликає значні порушення процесів обміну речовин, наприклад, обміну ліпідів (зниження рівня сфінгомієліну та ацетальфосфатидів у білій речовині головного та спинного мозку, порушення мієлінізації центральної нервової системи), хромо-протеїдів (падіння концентрації гемоглобіну) із затримкою дозрівання еритроцитів та зменшенням тривалості їх життя), синтезу еластину та колагену (ушкодження сполучної тканини, розрив аорти та серцевих судин), пуринового обміну (можливе підвищення активності ксантиноксидази, утворення сечової кислоти, підвищення активності уратоксидази); кислот (цитрату, малату, а-кетоглутарату, пірувату та інших).
Вивчення основної ролі металів в обміні речовин тваринного організму – важливий шлях досліджень впливу геохімічного середовища на тваринний організм. У біогеохімічних провінціях із надлишком молібдену дія цього металу пов'язана з посиленням синтезу ксантиноксидази та підвищенням її активності; у провінціях, збагачених бором, частково інгібуються протеїнази та амілази травного тракту; нестача йоду лімітує синтез гормонів щитовидної залози; недолік або надлишок кобальту, а також марганцю затримують використання йоду в синтезі трийодтиронінів та тироксину. Ці первинні впливу металів на процеси обміну речовин зумовлюють вторинні дисфункції багатьох біохімічних та фізіологічних процесів.
Оцінка можливості порушень обміну речовин чи взагалі появи біологічних ефектів під впливом геохімічних факторів середовища має ґрунтуватися на кількісних параметрах. Тому важливим завданням геохімічної екології є визначення меж концентрації хімічних елементів у ґрунтах, кормових рослинах, харчових раціонах, у межах яких забезпечується можливість нормального розвитку та життя тварин організмів, а також визначення порогових концентрацій, при яких порушується перебіг життєвих процесів.
Знання ґрунтових та харчових порогових концентрацій хімічних елементів дає можливість уявити географічну мінливість обміну речовин у тваринному організмі залежно від умов геохімічного середовища та може стати основою біогеохімічного районування.
Біосферу, яка відповідає території СРСР, ми розділили на регіони, названі біогеохімічними зонами. Вони обумовлюються єдністю ґрунтоутворювальних процесів, кліматичних факторів, біогенної міграції хімічних елементів та характером біологічних реакцій організмів на геохімічні та фізичні фактори середовища.
Зональні регіони біосфери поділяються на субрегіони - зональні біогеохімічні провінції, в яких комбінуються ознаки зон по концентрації та співвідношенню хімічних елементів, азональні, ознаки яких не відповідають характеристиці зон. Нами складено схематичну картку біогеохімічних зон і провінцій СРСР, де показано райони характерних змін обміну речовин та поширення низки ендемічних захворювань людини та тварин, а також райони загострення природного відбору під впливом нестачі або надлишку мікроелементів. Синтез окисних ферментів у тварин організмах за умов чорноземної зони виражений сильніше, ніж у нечорноземної зоні; у провінціях ендемічної атаксії в Дагестані, Узбекистані та в Актюбинському області спостерігається найбільш виражене гальмування синтезу окисних ферментів; вітамін Bi 2 депонується у тваринному організмі сильніше в чорноземній зоні порівняно з нечорноземною (особливо депонування вітаміну В, 2 у печінці та м'язах зменшено в районах піщаних ґрунтів); синтез йодних сполук щитовидної залози ослаблений у нечорноземній зоні та гірських зонах, а також заплаві річок інших зон; синтез ксантиноксидази посилено за умов молібденових провінцій Вірменії; уратоксидаза активніша у тварин у провінціях з підвищеним вмістом міді; у деяких випадках спостерігається різною мірою виражене інгібування протеїназ та амілаз у борних провінціях сухостепової, напівпустельної, пустельної біогеохімічної зон.
Для аналізу мінливості обміну речовин тварин необхідно враховувати як мінливість окремих особин, а й популяції загалом, що складається з індивідуумів одного виду, необхідно з'ясувати структуру популяції, використавши фізіологічну, біогеохімічну та морфологічну характеристики особин. У цьому індивідуальну мінливість слід як складову популяційної мінливості. Це дозволить усередині популяції розрізняти групи організмів із різною чутливістю до екстремальних геохімічних чинників. Цим створюється можливість спостерігати закономірності мінливості популяцій одного виду в різних умовах геохімічного середовища - при нестачі, надлишку або нормальному вмісті хімічних елементів у ґрунтах, водах, рослинах, кормах, харчових раціонах.
Мінливість порогової чутливості тварин до геохімічного середовища (мідь, молібден, бор, стронцій, уран та інші) ми широко досліджували. Внутрішньопопуляційна фізіологічна та біохімічна мінливість організмів визначає ступінь загострення природного відбору та ступінь адаптованості організму до екстремальних умов. Можна припустити, що у популяціях тварин накопичуються значні резерви потейціальної прихованої мінливості, зумовленої, очевидно, малими мутаціями генів та його рекомбінаціями. Такі резерви мінливості виявлені нами у ґрунтових мікроорганізмів, що живуть в умовах високої концентрації певних хімічних елементів (молібден, ванадій, бор, селен, кобальт). Внутрішньопопуляційна мінливість мікроорганізмів визначалася в нашій лабораторії за фізіологічною ознакою - пристосовуваність організмів окремих штамів, виділених з однієї колонії, до різних можливих природних концентрацій хімічних елементів (від мінімальних до максимальних). Наприклад, штами Bacillus megaterium з ґрунтів, багатих на уран, добре ростуть при високих концентраціях цього елемента і мало розвиваються при низьких його концентраціях. Навпаки, бактерії того ж виду, виділені з ґрунтів, бідних на уран, не можуть розвиватися при високих його концентраціях. Це загальне правило для різних видів мікроорганізмів та хімічних елементів. Такі ж закономірності зростання бактерій та низки актиноміцетів в умовах борних провінцій при порівнянні їх з такими ж видами, взятими з ґрунтів, бідних на бор. Серед досліджених штамів виявлені мутантні, що не підкоряються загальним правилам - виділені з ґрунтів, багатих на бор, але ростуть добре при будь-яких його концентраціях. Шляхом введення ДНК від мутантних форм і форм, адаптованих до високих концентрацій бору, в культурі мікроорганізмів, виділених з ґрунтів, бідних бором, можна отримати генетичну трансформацію пристосованості до бору - форми, добре ростуть при високих його концентраціях. Генетична трансформація була здійснена також у Bacillus megaterium, що живуть при низьких концентраціях селену, за допомогою ДНК, виділеної із форм селенової провінції. З непристосованих до селену були отримані форми, які добре розвиваються при високих концентраціях селену. При цьому необхідно вказати, що у цих бактерій із селенових провінцій виявлено захисний фермент - селен-редуктазу, який відновлює селеніти до елементарного селену, що не засвоюється. У форм цього ж виду, виділених із ґрунтів, бідних на селен, цей фермент не виявлений. Генетична трансформація призводить до появи ферменту у форм, які не синтезували його раніше.
За допомогою генетичної трансформації показано спадкову природу пристосованості мікроорганізмів до екстремальних умов хімічного середовища. Можна припускати, що в біогеохімічних провінціях, багатих або бідних на певні хімічні елементи, виникають мутації, індуковані екстремальними факторами. Відбувається збагачення генофонду популяції, що створює умови для загострення природного відбору та видоутворення, перетворення генетичної та екологічної структури популяції при екстремальних змінах умов геохімічного середовища.
Мінливість мікробних популяцій та їх генетичну природу можна вивчати (при вдалому виборі природних об'єктів) за допомогою звичайних мікробіологічних методів (експериментальна геохімічна екологія мікроорганізмів), тоді як для проведення подібних досліджень на рослинних та тваринних організмах необхідні умови фіто- та зоотронів, де можна регулювати та фізичні фактори середовища. Геохімічна екологія організмів повинна як спостерігати природні явища - вплив середовища проживання і співтовариств на організми, але має розвиватися як експериментальна наука (експериментальна геохімічна екологія).
Ми розглянули деякі питання геохімічної екології лише на рівні особин і населення. Геохімічна ж середовище діє організми всіх рівнях будови біосфери, лише на рівні регіонів біосфери (біогеохімічні зони) і субрегіонів (біогеохімічні провінції) біогеоценозів, популяцій, окремих особин.
Дослідження в галузі геохімічної екології, встановлення причинних залежностей неможливе без вивчення органів та тканин (концентрування хімічних елементів та вплив ступеня їх накопичення на проміжний обмін речовин, активність та синтез біологічно активних сполук, особливо ферментів). Такі дослідження, як і проведені на клітинному, субклітинному та молекулярному рівнях, є основою розуміння зв'язку організмів з геохімічним середовищем та адаптації організмів до геохімічних факторів середовища. Прикладом дослідження питань геохімічної екології на молекулярному рівні може бути аналіз впливу на організм різних концентрацій та співвідношень міді та молібдену. При високій концентрації у харчовому раціоні тварин молібдену та низької міді індукується молібденом синтез ксантиноксидази, підвищується її активність та зростає утворення сечової кислоти. При наростанні в організмі вмісту сечової кислоти вона індукує синтез уратоксидази, ферменту, що викликає деградацію сечової кислоти. Взаємини між речовинами, що беруть участь у аналізованих формах пуринового обміну, дуже складні. При збільшенні у раціоні кількості міді та поступовому падінні рівня молібдену відбуваються цікаві адаптивні зміни ксантиноксидази. При дефіциті молібдену та підвищеному вмісті міді її активність все ж таки зберігається. Нашими дослідженнями показано, що ксантиноксидаза молока за цих умов може збагачуватись міддю у 3,5-5,5 рази, втрачаючи молібден. Синтез уратоксидази у випадках індукується як сечової кислотою, а й міддю, вміст якої підвищено.
У цих експериментах при дії на організм різних співвідношень міді та молібдену може бути досягнутий високий оптимум активності ксантиноксидази, що не відповідає фізіологічному оптимуму, що спостерігається при співвідношенні у раціоні міді та молібдену 1:4.
Ця раніше невідома форма молекулярної адаптації процесів в організмі щура, очевидно, є фенотипічним проявом генної функції ферменту.
При екологічних дослідженнях, в такий спосіб, можна підійти до аналізу процесу еволюції організмів - мінливості, пристосовності, утворення нових таксономічних одиниць, природного відбору та його генетичних основ. Для вивчення питань еволюції відкриває шлях геохімічна екологія організмів та його популяцій в екстремальних умовах середовища - при надмірному чи недостатньому вмісті мікроелементів у біосфері.
Надіслати свою гарну роботу до бази знань просто. Використовуйте форму нижче
Студенти, аспіранти, молоді вчені, які використовують базу знань у своєму навчанні та роботі, будуть вам дуже вдячні.
Курсова робота
на тему:
« Біохімічні шляхи у дослідженні механізмів психічних та нервових хвороб»
ВСТУП
Патологічні стани центральної нервової системи численні, різноманітні та надзвичайно складні за механізмом виникнення та розвитку. У цій роботі будуть показані тільки шляхи, на яких вчені-біохіміки досягли певних успіхів у пізнанні окремих елементів патологічних процесів, що лежать в основі низки хвороб та хворобливих станів центральної нервової системи.
1. ХІМІЧНІ ФАКТОРИ ВНУТРІШНЬОГО ПІДКРІПЛЕННЯПРИ НАРКОМАНІЯХ
У примітивно влаштованих організмів ланцюжок, реакцій у центральній нервовій системі, що починається під дією тієї чи іншої мотивації, наприклад голоду, інстинкту продовження роду і т.п., має мінімальну кількість проміжних ланок і завершується безпосереднім досягненням або недосягненням мети. Досягнення кінцевої мети є найпростішим підкріпленням, фактором, що винагороджує. У відносно високорозвинених організмів процес досягнення мети може бути поділений на більшу кількість етапів. Завершення проміжного етапу не винагороджується кінцевим результатом і, як встановлено, існує так звана система внутрішнього підкріплення.Наприклад, виконуючи певну роботу, сучасна людина, як правило, не отримує після її завершення харчової винагороди, але отримує гроші, відчуваючи при цьому певне задоволення та впевненість у можливості за допомогою грошей реалізувати мету – придбання та споживання їжі. У центральній нервовій системі почуття задоволення може бути забезпечене за допомогою низки гуморальних факторів, до яких насамперед відносяться нейропептиди - деякі з опіоїдів, нейротензин та ін.
Найпростіший експеримент, що дозволяє у дослідах на тваринах виявити ці гуморальні чинники, полягає у наданні їм можливості вибору речовин, що викликають приємні відчуття при самовведенні в шлуночок мозку. Білий щур, наприклад, з вживленими в мозок канюлями, що має можливість натисканням тієї чи іншої педалі ін'єкувати собі розчин однієї з випробовуваних речовин, досить швидко переходить від безладного натискання різних педалей до свідомо кращого самовведення досить вузького кола речовин. Існують і складніші форми такого роду експериментів.
В результаті, до категорії речовин, що віддають перевагу - ймовірних внутрішніх факторів підкріплення - зараз відносять деякі з опіоїдних нейропептидів - р. -Еідорфіні еїке-фаліни, а також нейротензин.Одночасно виявляються і нейропептиди, які мають протилежну дію: вазопресині, мабуть, меланостатині тіроліберін.
Вводячи собі ззовні нейропептиди винагороди, тварина обходиться без нормального механізму, необхідного у природних умовах забезпечення шляху до мети, завершення якого винагороджується утворенням внутрішнього хімічного сигналу - чинника внутрішнього задоволення.
Багато дослідників розглядають ці експерименти як модель наркоманії. Істота наркоманії полягає, з цього погляду, у підміні зовнішнім хімічним агентом природної внутрішньої хімічної винагороди. Така підміна при доступності хімічного еквівалента внутрішнього фактора не вимагає цілеспрямованої праці та інших процесів для того, щоб досягти стану задоволення, насолоди тощо. Екзогенні опіати - морфінта його аналоги – є еквівалентами внутрішніх опіоїдів, взаємодіючи з тими ж класами рецепторів головного мозку, що й опіоїдні нейропептиди. Характерно, що найбільш апробованим засобом зняття абстинентних станів у наркоманів є специфічний блокатор опіатних рецепторів. налоксон,а також його аналоги.
Механізми наркоманій, викликаних опіатами, знаходять таким чином тлумачення, яке можна віднести до категорії досить обґрунтованої гіпотези. Складніше ситуація з низкою інших наркотиків - кокаїном, каніабіноїдами, ЛСД, мецкалінамита інших. Щодо деяких їх розглядаються гіпотези, аналогічні викладеної вище. Зокрема, катехол аміни,особливо норадреналін,і серотонін,у певних зонах мозку беруть участь у процесах внутрішнього підкріплення. Про це свідчить наявність зон, розташованих по ходу катехоламінергічних та серотонінергічних шляхів, подразнення яких викликає відчуття задоволення, задоволення тощо. Багато із згаданих вище наркотинів відомі як агенти, що втручаються в катехоламінергічну та серотонінергічну нейротрансмісію. Тому, хоч і з меншою мірою доказовості, ніж у випадку опіатних наркотиків, гіпотеза про наркоманію, як заміну факторів внутрішнього підкріплення, є правомочною і для багатьох неопіатних сполук. Слід також мати на увазі, що механізми та хімічні фактори внутрішнього підкріплення відомі лише частково.
У рамках викладеної гіпотези намагалися укласти й дані про механізми алкоголізму. Простежуються складні зв'язки між розвитком алкоголізму, рівнями опіоїдних пептидів, непептидних факторів, подібних до опіатів, катехоламінів, серотоніну та інших гуморальних регуляторів. Складність цих зв'язків така, що, крім ролі чинників внутрішнього підкріплення, доцільно розглянути ряд даних про механізми алкоголізму особливо.
2. АЦЕТАЛЬДЕГІД, НЕПЕПТИДНІ ТА ПЕПТИДНІ ОПІОЇДИ ТА АЛКОГОЛІЗМ
Перші дослідження біохімічних механізмів алкоголізмупризвели до встановлення трьох важливих фактів. По-перше, етанол є мембранрогітним агентом і може в концентраціях, що викликають сп'яніння, змінювати стан рецепторів та багатьох ензимів, інкорпорованих у мембрану. По-друге, широко представлений в організмі, особливо в печінці, фермент - алкогольдегідрогеназу,що бере участь у метаболізмі багатьох регуляторних сполук, входить у контакт із етанолом, що надходить ззовні, і як би відволікається від ряду нормальних функцій, що, у свою чергу, веде до відхилень у синтезі ряду регуляторів. По-третє, нарешті, алкогольдегідрогеназу швидко перетворює частину надходить етанолу в ацетальдегід; останній може бути джерелом утворення низки біоактивних факторів і, крім того, його пряма дія на мозок викликає неприємні відчуття синдром похмілля; далі ацетальдегід поступово окислюється мітохондріальною ацетальдегіддегідрогеназоюі ацетат, що утворюється, може служити для синтезу жирних кислот і т.п. Відчуття, що викликаються ацетальдегідом, з'явилися відправною точкою для створення ряду протиалкогольних засобів, що пригнічують ацетальдегіддегідрогеназу, що підвищують тим самим рівень, ацетальдегіду і прискорюють, в результаті, розвиток тяжкого стану після прийому навіть невеликих доз алкоголю. У практику увійшов, зокрема, такий інгібітор цього ферменту, як тетурам,систематичне запровадження якого приносить певну користь у лікуванні алкоголізму. Особливо перспективними виявилися впливи, що дозволяють тривалий час змінювати активність двох зазначених головних ферментів метаболізму етанолу. В експериментах на щурах-алкоголіках ефективною виявилася індукція аутоантитіл, що пов'язують ці ферменти.
а взаємодіючи з серотоніном, - метил-тетрагідро--карбо-лін:
Ці сполуки мають деяку структурну подібність із морфіном.
Адетальдегід здатний також гальмувати один з етапів катаболізму дофаміну - його окисне дезамінування - так що накопичується проміжний продукт - 3,4 - діоксі-фенілацетатальдегід.Останній, взаємодіючи знов-таки з дофаміном, утворює тетрагідропапаверолін,здатний, у свою чергу, перетворюватися на сполуки, що все більш наближаються за структурою до морфіну, у тому числі - норморфін:
В останні роки простежено метаболічні шляхи, що ведуть до утворення в організмі ссавців навіть кодеїну та морфіну, хоча й у дуже малих кількостях. Більше того, зараз можна вважати, що в мікроконцентраціях багато сполук цього ряду постійно представлені в мозку. Однак введення ззовні етанолу та утворення з нього ацетальдегіду різко підвищує рівень морфіноподібних сполук.
Встановлено здатність сальсолінолу та інших ендогенних аналогів морфіну, що утворюються за участю ацетальдегіду, служити як агоністами, так і блокаторами опіоїдних рецепторів залежно від концентрації та інших умов. Наслідки такої взаємодії можуть полягати, по-перше, у підміні ендогенних факторів винагороди і, по-друге, якщо концентрація сальсолінолу в організмі алкоголіка постійно підвищена, то блокада рецепторів щодо власних ендогенних, найбільш адекватних факторів винагороди може викликати постійне почуття незадоволеності та спонукання для пошуку наркотичних засобів.
Поки що важко віддати перевагу одній з цих можливостей, але утворення сальсолінолу та подібних до нього непептидних морфіноподібних сполук при алкоголізмі вказує на ймовірний зв'язок опіоїдної системи з механізмом алкоголізму. До цієї ж думки наводить той факт, що класичний блокатор опіатних рецепторів. налоксонвиявився корисним також під час лікування алкоголізму.
Нарешті, показово, що більшість алкоголіків зростає рівень антитіл до морфиноподобным сполукам. Тому зрозуміла наполегливість, з якою сучасні дослідники після відкриття опіоїдних пептидів шукають кореляції між рівнем останніх, а також станом опіоїдних рецепторів, з одного боку, і глибиною і фазою алкоголізму, з іншого боку. Знайдені зараз кореляції, що підтверджують більш менш значну участь системи опіоїдів у механізмах алкоголізму, свідчать про досить складні відносини, часом суперечливі. Так, наприклад, показано менший зміст метенкефалінуу мозку схильних до алкоголю тварин і менша концентрація в гіпоталамусі р -ендорфінуу тварин зі сформованим алкоголізмом та спадково схильних до алкоголізму.
Встановлено також, хоч і не на всіх експериментальних моделях, що введення таким тваринам етанолу підвищує рівень метенкефаліну та р-ендорфіну.
Можна намагатися тлумачити ці дані те, що знижені рівні ендогенних опіоїдів у мозку зумовлюють потяг до етанолу як до чинника, що веде до освіти опіоїдів мозку, тобто. до нормалізації гуморальних систем винагороди З цим узгоджуються феномени зняття абстиненції та деякого зниження потягу до алкоголю під час введення ззовні опіоїдних нейропептидів, а також деяких інгібіторів протеолітичного розпаду опіоїдних пептидів в організмі. Однак ряд експериментальних даних важко узгодити з таким тлумаченням. Так, на відміну від метенкефаліну вміст лейенкефаліну в мозку схильних до алкоголізму тварин підвищено.
Суперечливі дані про зміни рівня р-ендорфіну в плазмі та цереброспінальній рідині при введенні етанолу. Джерелом непорозумінь є також те, що при алкоголізмі можливі, і в ряді робіт зареєстровані, не тільки зміни рівня ендогенних опіоїдів, але й стану їх рецепторів через згадану мембранотропність етанолу. Є дані про зниження спорідненості енкефалінів до їх рецепторів під дією етанолу. Отже, для суворого обліку ролі ендогенних опіоїдів при алкоголізмі необхідний спільний розгляд даних про рівень опіоїдів та стан їх рецепторів. Таких даних поки що недостатньо.
Багатий, хоча знов-таки неоднозначний, експериментальний матеріал зібраний про роль катехоламінів та серотоніну у розвитку алкоголізму. Тут простежується чітка залежність дії етанолу від етапу, фази розвитку алкоголізму.
Одноразовий прийом етанолу викликає спочатку посилений викид катехоламінів - дофаміну та норадреналіну, зумовлений, ймовірно, мембранотропною дією алкоголю на пресинаптичні рецептори. Катехоламіни, що секретуються, входять до числа факторів внутрішньої винагороди, що викликають ейфоричний стан. Після викиду катехоламінів спрацьовує система зворотної регуляції, яка не просто нормалізує стан системи, а створює тимчасовий дефіцит катехоламінів у синаптичній щілині та рідинах організму. Можливо, це є одним із спонукаючих факторів до повторення прийому етанолу.
При розвитку хронічного алкоголізму стан зниженого виходу, посиленої деградації та підвищеного зворотного захоплення катехоламінів як би закріплюється, створюючи, як вважають, постійний механізм, що спонукає до частого прийому етанолу для тимчасової корекції цих порушень. Повне припинення прийому алкоголю на стадії розвиненого хронічного алкоголізму веде до екстреної мобілізації всіх існуючих механізмів синтезу, викиду та збереження катехоламінів у синаптичній щілині. Розрегульована на попередніх стадіях система спрацьовує так, що відбувається не нормалізація рівня катехоламінів, а надмірне зростання їхньої концентрації, зокрема дофаміну. Вони беруть участь у розвитку абстиненції.
Подібною є, загалом, і динаміка змін виходу та перетворень серотоніну. Збочення його викиду можуть бути пов'язані з ейфорією та галюцинаціями. Разом з тим роль серотонінергічної системи видається поки неоднозначною - так, з одного боку, ряд стимуляторів синтезу та виходу серотоніну, блокатори зворотного захоплення та багато агоністи пригнічують потяг до етанолу. У той же час селективні антагоністи 5НТ 3 -рецепторів та такі інгібітори синтезу серотоніну, як р-хлорфенілаланін, також пригнічують споживання етанолу. Активна імунізація тварин проти серотоніну веде до зниження його рівня в плазмі крові та в мозку та до придушення потягу до алкоголю експериментальних тварин. Очевидно, участь серотонинергической системи у механізмах потягу алкоголю дуже тісно пов'язані з типом рецепторів серотоніну, і навіть локалізацією як рецепторів, і місць синтезу цього медіатора.
У механізми наркоманій та алкоголізму залучено також головну гальмівну систему мозку - ГАМК-ергічну. Пригнічення цієї системи дозволяє зрозуміти стійкість патологічного потягу.
Характерно, що дія на потяг до алкоголю низки нейропептидів більш-менш корелює з їх участю у розвитку чи придушенні стресових станів. Взагалі відомо, що стрес сам собою стимулює потяг до алкоголю. Пептид дельтасна, що виявився сильним протистресовим агентом, достовірно знижує споживання етанолу експериментальними тваринами при його систематичному введенні. У схильних до алкоголю білих щурів його вміст у плазмі крові та стріатумі знижено.
Загалом представлені результати досліджень у галузі нейрохімічних механізмів алкоголізму не утворюють поки що єдиної стрункої картини, але свідчать про відносну близькість часу, коли вона сформується.
3. СТРАХ, ФОБІЇ,р-КАРБОЛІНИ, ЕНДОЗЕПИНИ ТА ХОЛЕЦИСТОКІНІН_4
Порівняно давно фармакологи створили новий клас транквілізаторів. бензодіазепіни,що увійшли зараз у широку медичну практику. Потім було виявлено рецептори цих сполук у головному мозку. Оскільки були відомі внутрішні ліганди цих рецепторів, їх позначили як рецептори діазепаму. Далі виявилося, що ці рецептори є частиною рецепторів гамма-аміномасляної кислоти або самостійним рецептором, міцно пов'язаним з рецептором ГАМК. Нарешті вдалося виділити частину ендогенних лігандів цих рецепторів: по-перше, великий пептид - евдозепін,що складається приблизно з сотні амінокислотних залишків, його активні фрагменти - малі 18 - і 6_членні пептиди і, по-друге, непептидні сполуки - похіднітак званих -карболінів.Прикладом останніх є метилтетрагідро-карболін. Активні фрагменти ендозепінів мають структуру: QATVGDVNTDRPGLLDLK та GLLDLK.
Ці сполуки впливають на поведінку тварин, зворотне дії ГАМК та її аналогів. Вони викликають занепокоєння, прояви страху та у дослідах на гризунах проконфліктну поведінку. У документовано гостре занепокоєння, панічний стан людей, яким вводили одне з похідних -карболіна.
Що стосується транквілізаторів - бензодіазепінів, з яких почався цей цикл досліджень, то вони виявилися блокаторами рецепторів ендозепінів, що пригнічують їх взаємодію з ендогенними факторами страху, занепокоєння та проконфліктної поведінки.
В останні роки увагу нейрохіміків і психіатрів привернув ще один пептид, що викликає занепокоєння, страх і панічну поведінку як у людей, так і у тварин, - найменший із виявлених у мозку С-конпевих фрагментів холецистокініну - ХЦК_4. Його дія на поведінку опосередкована стимуляцією деяких відділів дофамінергічної системи через спеціальні рецептори ХЦК В. Вже синтезовано антагоністи ХЦК_4, за допомогою яких вдається знизити рівень тривожності та панічних реакцій як у дослідах на тваринах, так і в перших клінічних дослідженнях.
Для розуміння біохімічних механізмів низки розладів психіки значення цих відкриттів дуже велике. Багато психічних розладів супроводжуються нав'язливими страхами, фобіями, вкрай неспокійною та конфліктною поведінкою. Вони характерні, зокрема, для пізніх стадій алкоголізму, деяких проявів шизофренії та інших.
4. ДОФАМІН І ПАРКІНСОНІЗМ
Розкриття біохімічних процесів, що лежать в основі хвороби Паркінсона- глибокого порушення стереотипної рухової активності, її координації та ініціації,- стало одним із перших яскравих досягнень патологічної нейрохімії.
Синдром хвороби вдалося відтворити в експериментах на тваринах, вводячи їм 6 _оксидофамін. Цей аналог дофаміну проникає у везикули нервових закінчень, призначені для накопичення та викиду катехоламінів, конкурує з останніми за включення у везикули і, зрештою, пригнічує катехоламінергічну трансмісію.Цей процес іноді називають хімічною десимпатизацією,маючи на увазі особливу роль катехоламінів у симпатичній нервовій системі. Однак ця назва неточна, бо катехоламіни широко поширені та функціонують у багатьох інших відділах нервової системи.
Дофамінергічні нейрони стріатуму, частини хвостатого ядра і особливо чорної субстанції, які є основними центральними організаторами стереотипної рухової активності, виявилися високочутливими до такої дії 6-оксидофаміну. В результаті вперше вдалося, вводячи речовину певної біохімічної спрямованості дії, викликати таке специфічне захворювання, як паркінсонізм.
В останні роки з'явилася можливість ще точнішого визначення нейронів, пошкодження яких достатньо для виникнення паркінсонізму. Синтетичний нейротоксин метилфенілтетрапедропіридинвибірково зв'язується з меланінсодержащими нейронами чорної субстанції, викликаючи їх депігментацію та паркінсонічний синдром. Відзначено також суттєве зниження вмісту у чорній речовині метенкефалннаі холецистокінінаі виявлено, нарешті, дефіцит одного з гліальних білків, що виконують тропічні функції по відношенню до нейронів, що синтезують глутамін.
Участь дофамінергічних систем у паркінсонічному синдромі передбачає можливість полегшення синдрому введенням у мозок дофаміну. Оскільки дофамін не проходить гематоенцефалічний бар'єр, скористалися для введення хворим його найближчим попередником. діоксифенілаланіном.Він суттєво полегшує стан паркінсоніків. Слід, однак, підкреслити, що тривале введення хворим великих доз діоксифенілаланіну, що значно посилює синтез дофаміну у всіх відділах мозку, може вести до появи симптомів, подібних до іншого психічного захворювання - шизофренія, одним з фізіологічних і біохімічних проявів якого є саме гіперактивність дофа.
Новий шлях лікування паркінсонізму полягає у пересадках клітин або ділянок тканини здорового мозку з мезенцефалону людських плодів, здатних продукувати дофамін, у певні ділянки мозку хворого. Зауважимо, що особливості імунологічного статусу мозку значно полегшують такі пересадки. Результати перших серій таких пересадок обнадіюють. Зараз накопичується досвід тривалого спостереження наслідків цих операцій і розробляються культури клітин мезенцефалону для того, щоб відмовитися від використання матеріалів, що отримуються при абортах.
5. ШИЗОФРЕНІЯ, КАТЕХОЛОМІНИ ТА ВНУТРІШНІ НЕЙРОЛЕПТИКИ
Практично повна відмова сучасної психіатрії від палат для буйних психічних хворих пояснюється двома причинами, різними зовні, але подібними насправді. Перша полягає в хірургічному втручанні - перерізанні катехоламінергічних шляхів, що йдуть до лобової кори від таламуса, ретикулярної формації, чорної субстранції та деяких інших відділів мозку. Досить ефективний зі зняттям агресивних проявів шизофренії, це, тим щонайменше, пов'язані з певної деградацією розумових здібностей і нині поступилося місце фармакологічним впливам. Останні полягають у придушенні рецепції та/або секреції катехоламінів, особливо дофаміну, такими сполуками, як галоперидол, трициклічні нейролептикита ін. Строго кажучи, ці агенти не так лікують хворих від шизофренії, скільки пригнічують її прояв: агресивну поведінку, галюцинації, стереотипну рухову активність тощо.
Ефективність зазначених засобів наближає до розуміння механізмів хвороби, що включають глибоке збочення та патологічне посилення у певних відділах, мозку шизофреніків катехоламінергічної, особливо дофал. інергічної, трансмісії. Встановлено 4-5_кратне підвищення густини рецепторів дофаміну D 4 . Показово, що одне з кращих антипсихотичних ліків - клозапін - має найбільшу спорідненість саме до рецепторів D 4 . Виявлено також значно підвищений рівень дофаміну у скроневій частці головного мозку, особливо у лівій мигдалині. Відзначено й низку морфологічних змін у тих самих відділах - збільшення обсягу бічних шлуночків, витончення парагиппокампальной кори та інших.
Суперечливими, на відміну закономірних змін змісту дофаміну, є відомості про вміст норадреналіну в різних відділах мозку шизофреніка. Щодо відтворення лише дані про підвищення рівня норадреналіну в цереброспінальній рідині.
На особливу увагу заслуговують повідомлення про відтворення окремих проявів шизофренії при впливі агентів, що так чи інакше втручаються в стан катехоламінергічної та серотонінергічної систем. Так, аналог дофаміну - рослинний алкалоїд мецькалінвикликає галюцинації, у тому числі кольорові, що мають подібні елементи з шизофренічними.
Тварини, які отримали високі дози фенаміну,після періоду збудження виявляють монотонну, стереотипну рухову активність, що нагадує таку у шизофреніків.
Галюцинаторні явища, які спостерігаються при введенні деяких аналогів серотоніну, наприклад діетиламіду лізергінової кислоти, також заслуговують на увагу з точки зору можливої ролі при шизофренії не тільки збочень катехоламінергічної, а й серотонінергічної трансмісії.
Картина патологічного, незбалансованого посилення при шизофренії катехоламінергічних та, можливо, серотонінергічних систем у певних ділянках мозку добре узгоджується і з даними про зміну набору та активності моноамінооксидаз, що розщеплюють відповідні нейромедіатори після виходу з нервового закінчення. З чотирьох форм МАО, виявлених у мозку людини, - I, II, II і III - у шизофреніків відсутня III і суттєво модифікована II форма.
Нарешті, до механізмів шизофренії залучено ще одну нейромедіаторну систему - глутаматергічну. Відзначено її значне ослаблення у передній корі. Нагадаємо, що з цією системою особливо тісно пов'язана низка вищих функцій мозку. Такий антагоніст глутамату, як пенциклідин, імітує деякі симптоми шизофренії на тваринах. Наразі вивчається можливість застосування агоністів глутаматергічних рецепторів для терапії шизофренії.
Менше даних накопичено поки щодо змін пептидної регуляції при шизофренії.
У нервових закінченнях, зазвичай, зустрічаються ті чи інші асоціації класичних яюйромедіаторів з нейропептидами. Зокрема, дофамінергічні закінчення особливо часто містять холецистокінін _8, вазоактивний інтестинальний пептиді, рідше, соматостатині речовина Р.У норадренренгічних закінченнях частими супутниками норадреналіну є нейропептидиYі енкефаліни.Відомо також, що холецистокинин_8 і, з меншою визначеністю, соматостатин гальмують дофамінергічну передачу. Аналогічні дані є щодо дії енкефалінів на норадренергічну трансмісію. Активатором же норадренергічної
передачі є нейропептид Y, а дофамінергічною, знов-таки з меншою визначеністю, - вазоактивний інтестинальний пептид. Тому суттєве значення мають дані про зниження вмісту холецистокініна_8 в гіпокампі та мигдалику мозку шизофреніків. Знижено також вміст у гіпокампі соматостатину, а в мигдалині - метенкефаліну та речовини Р, проте концентрація вазоактивного інтестинального пептиду в мигдалині підвищена.
З усією цією групою даних асоціюються відомості про нейролептичну дію холецистокініна_8 та спорідненого йому пептиду. церулеїну,а також деяких ендорфінів.Воно проявляється у тестах, розроблених у свій час для відбору ліків-нейролептиків: по здатності викликати каталепсію, по дії на агресивну поведінку, придушення ефектів фенаміну та ін. Зниження ними дофамінергічної трансмісії показано і в експериментах на клітинному рівні. Все це послужило основою для найменування холецистокініна_8, а також дезтирозил-і дезенкефалін-ендорфінів ендогенними нейролептиками. На жаль, спроби використовувати їх як лікувальні засоби для усунення шизофренічного синдрому у людей поки не дали відчутних результатів. Можна говорити поки що лише про позитивні тенденції, зокрема про перспективність спільного застосування холецистокініну або церулеїну із звичайними нейролептиками.
Найбільш перспективними видаються дослідження, засновані на даних про здатність короткого кінцевого тетрапептиду холецистокініна - ХЦК_4 - викликати тривогу, страх і паніки. Дія ХЦК_4 знову ж таки пов'язана з активацією частини дофамінергічної системи. Тому на синтезі та випробуваннях антагоністів ХЦК_4 засновані надії на створення нового класу лікарських речовин для лікування деяких форм шизофренії.
6. КАТЕХОЛОМІНИ, НЕЙРОПЕПТИДИ І ДЕПРЕСИВНІ СТАНИ
Пригнічення катехоламінергічної та серотонінергічної систем пов'язане з депресивним станом. Однак картина придушення катехоламінергічної трансмісії при депресіях не є простим зверненням того, що спостерігається, наприклад, при агресивних формах шизофренії, а механізми депресій не зводяться тільки до порушень класичних медіаторних систем.
Нагромадилося чимало даних про суттєве зниження норадренергічної трансмісії при депресіях. Найменшою мірою знижується активність серотонінергічної системи. Ще менше зрушень у дофамінергічній трансмісії. Відсутні значні зміни в інших класичних нейромедіаторних системах. У згоді з цими даними є ефективність зняття депресії великою групою лікарських засобів. іміпраміном, амітршгіліном, інказаномта багатьма іншими, дія яких полягає або у гальмуванні зворотного захоплення норадреналіну та серотоніну, або в інгібуванні їх розщеплення моноамінооксидазами. Зауважимо лише, що такий механізм дії антидепресантів лише на початкових етапах застосування. При тривалому їх введенні включаються складні зміни у рецепції медіаторів. При введенні речовин, що знижують рівень катехоламінів, може виникнути депресія. Такою речовиною виявився резерпін,застосовується під час лікування гіпертонії.
Таким чином, значущість зниження активності катехоламінергічної системи при розвитку депресій очевидна; однак це зниження не є простою протилежністю до тих змін, які описані для шизофренії. При шизофренії – переважне посилення дофамінергічної трансмісії, при депресії – переважне пригнічення норадренергічної та серотонінергічної. Вважають також, що у характерному для депресій придушенні психічної та рухової активності бере участь також дисбаланс між збудливою глутаматергічною та гальмівною ГАМК-ергічною системами, що відзначається як при депресіях, так і при шизофренії. Нарешті, у мозку хворих на депресії виявлено значні, зміни щільності опіатних рецепторів. Це свідчить про можливі відхилення у системі «внутрішнього винагороди».
7. СУДНОВІ СТАНИ, ЕПІЛЕПСІЯ,ГЛУТАМАТНІ РЕЦЕПТОРИ ТА ЇХ АНТАГОНІСТИ
Біохімічні механізми судомних станів і особливо такої хвороби, як епілепсія, принципово відрізняються від описаних вище для шизофренії, депресій, наркоманій та алкоголізму. Важлива роль індукції синдрому епілепсії належить глутаминергической системі. Очевидно також значення ГАМК-ергічної та ендозепамергічної систем.
З глутамінергічною трансмісією тісно пов'язана не лише можливість індукції судомних станів, а й низка вищих функцій ЦНС, таких, наприклад, як пам'ять. У цьому одна з причин того, що епілепсія не зводиться до судомного синдрому і пов'язана з складними змінами психіки.
Рецептори глутамінової кислоти – утворення складні та неоднорідні. Характерно, що розкриття їх структури та різноманітності було прискорено виявленням речовин, які іноді називають збуджуючими нейротоксинами. До них належить каїнова кислота, квісквалева кислотата ряд інших сполук, багато з яких мають спільні з глутаматом елементи структури. Сам собою глутамат при інтрацеребральному введенні у певні зони мозку може викликати напади судом. Однак каїнат і квісквалат виявилися особливо потужними індукторами судом і, більше того, агентами, здатними специфічно руйнувати нейрони, що несуть глутаматні рецептори.
На глутаматних рецепторах виявлено ділянки зв'язування барбітуратів- Агентів, які гальмують їх функцію і мають відповідно протисудомну активність. Один з найпотужніших і найспецифічніших блокаторів NMDA_глутаматних рецепторів - 2 _амино_7_фосфоногептанова кислота- запобігає нападам епілепсії у експериментальних тварин. Все це змушує вважати зміни глутамінергічної трансмісії одними із вузлових у патогенезі епілептиморфних судомних станів.
Підтримкою цієї гіпотези служить виявлення в плазмі крові епілептиків значно підвищених рівнів антитіл до білків глутаматного рецептора. Це використовується для діагностики прихованих форм епілепсії та оцінки тяжкості захворювання. Подібне явище відображає, мабуть, зниження функцій гематоенцефалічного бар'єру при розвитку епілепсії, що супроводжується виходом у периферичний кровотік певних кількостей білків рецептора та їх фрагментів, і далі утворенням антитіл до них. Можливо, сам патогенез епілепсії подібний до патогенезу аутоімунних хвороб мозку, при яких аутоантитіла до білків мозку служать основним ушкоджуючим фактором. У тому, що остання гіпотеза є правомочною, переконують, по-перше, дані про підвищений рівень у плазмі крові епілептиків не тільки антитіл до білків глутаматного рецептора, але й до інших білків та ліпідів синаптичних мембран мозку, наприклад до білка S_100, а також результати експериментів. з введенням у мозок антитіл, отриманих до різних білкових та ліпідних фракцій мозку. Епілептиформні процеси виникали, зокрема, при ін'єкції антитіл до деяких фракцій гангліозидів.
Інша система,зв'язок якої з судомними станами та епілепсією дуже ймовірний, - ГАМК-ергічна.
Гальмівні функціїГАМК-ергічної системи мають більш універсальний, менш специфічний характер, ніж функції збуджуючих нейромедіаторних систем. Це відображає, зокрема, той факт, що частка ГАМК-ергічних терміналей у мозку є найбільшою. Зниження судомної готовності та полегшення судомних станів встановлено при центральному введенні ГАМК, а також при периферичному введенні його аналогів, здатних проходити через гематоенцефалічний бар'єр. Такий самий ефект сполук, що гальмують розпад, стимулюють синтез і зворотне захоплення ГАМК: вальпроату натрію, а також прогабіда,у -ацетилен-ТАМКта ін.
На рецепторах ГАМК виявлено барбітурат-зв'язуючу ділянку. У цьому випадку, на відміну від аналогічної ділянки на глутаматних рецепторах, барбітурати посилюють ефект основного нейромедіатора.
Таким чином, барбітурати виявляють протисудомну дію, змінюючи стан двох категорій рецепторів: глутаматних, пригнічуючи збуджуючу дію їх лігандів, та ГАМКд-рецепторів, стимулюючи їхню гальмівну дію.
Ще більше підтверджує уявлення про роль ГАМК-ергічної системи у запобіганні та усуненні судомних станів той факт, що придушення синтезу ГАМК, навпаки, провокує судоми. Яскравим прикладом є судоми людей при авітамінозі В 6 . Вітамін В6 служить попередникам піридоксаль_5_фосфата, який, у свою чергу, є кофактором глутаматдекарбоксилази,каталізує останню стадію синтезу ГАМК. Провокують судоми та такі інгібітори цього ферменту, як аллілгліцин, гідразидита ін Судомні напади викликає і один з отрут грибів - пікротоксинін,що теж зв'язується з ГАМК-рецепторами і пригнічує їх активність. З ГАМК-рецепторами пов'язані ділянки зв'язування ендозепінів - пептидів, що викликають збудження, страх і проконфліктну поведінку. Вони знижують активність ГАМК-рецепторів. Зрозуміло тому, що блокатори рецепторів ендозепінів – бензодіазепінові транквілізатори – виявилися агентами, які полегшують епілептичні процеси.
У цьому зв'язку, хоча матеріал, що викладається, стосується природи епілепсії та епілептичних судом, слід зазначити і дані про патогенез судом, що викликаються такими отрутами, як стрихніні правцевий токсин.Стрихнін блокує рецептори гліцину, другого за значимістю після ГАМК гальмівного медіатора центральної нервової системи, що функціонує переважно у спинному мозку. Дія ж правцевого токсину спрямована переважно на блокаду виходу ГАМК із нервових закінчень у головному мозку, що веде до блокади гальмівних впливів на мотонейрони. Ці дані знову-таки наголошують на ролі порушень систем гальмівних нейромедіаторів.
Була б, однак, помилковою спробою звести патогенез епілепсії та ряду інших судомних станів лише до змін глутамінергічної системи або її балансу з ГАМК-ергічною системою. Яскравою ілюстрацією складності проблеми та неприпустимості поспішного формування гіпотез у цій галузі є, наприклад, дані про здатність опіоїдного пептиду - метенкефаліну - викликати судоми при аплікації в деяких ділянках мозку - поза зонами, де він виявляє аналітичну дію, типову для опіоїдів. Ці дані послужили також основою припущень, що епілептичний напад - результат надмірно високого викиду енкефаліну з опіоїдергічних терміналей з поширенням його на зони мозку, де він здатний індукувати судоми. Передепілептичний стан - аура - тлумачилося при цьому як ейфоригенна дія опіоїду. На жаль, вся відома сукупність даних про механізми епілепсії не дозволяє прийняти таке просте тлумачення. Тут особливо важливо поєднане дослідження біохімічних та фізіологічних механізмів епілепсії.
Особливої уваги заслуговують останнім часом дані про роль пептидергічних систем при епілепсії і про існування ендогенних пептидних лігандів глутаматних рецепторів.
Патогенез судомних станів і епілепсії на відміну багатьох інших психічних хвороб тісно пов'язані з змінами енергетичних процесів у нейронах, причому у тих, які входять у патологічні епілептогенні осередки.
Відзначено зміни метаболічної структури нейронів та їх мітохондрій: підвищення проникності мембран для К+ та Na+, що корелює з підвищенням чутливості білків мембран до протеїназ, зниження синтезу АТФ та ін. Аномальні локальні зміни концентрацій К+ та Na+, а також NH4+, що виділяється при епілептичному нападі внаслідок посилення реакцій дезамідинування, можуть викликати або полегшувати деполяризацію постсинаптичних зон, знижувати поріг збудливості та провокувати судомний напад.
З підтримкою енергетичного статусу мозку тісно пов'язана система аденозинових рецепторів Aj. Ці рецептори виконують ряд гальмівних енергозберігаючих функцій, викликаючи седативні та протисудомні ефекти. Включення їх здійснюється аденозином та аденозинмонофосфатом - кінцевим продуктом утилізації таких макроергів, як АТФ та АМФ. Показано зміни рівня цих сполук при епілепсії та вивчається можливість створення на цій основі нових протиепілептичних засобів.
Наразі важко остаточно вирішити питання про первинність порушень у медіаторних, рецепторних чи енергетичних системах у патогенезі епілепсії. Безсумнівна, однак, їхня взаємодія. На це вказують глибокі локальні зміни енергетичних систем при ін'єкції каїнової кислоти саме у патологічно змінених ділянках епілептичного мозку.
8. ХОЛІНЕРГІЧНІ СИСТЕМИ МОЗКУ,р -АМІЛОЇД, НЕЙРОПЕПТИДИ І СЕНІЛЬНІ ДЕМЕНЦІЇ
Різновид старечого недоумства, так званий сенільна деменція Альтцгеймерівського типу,супроводжується насамперед дегенерацією великої кількості підкіркових нейронів, холінергічні терміналі яких широко поширені у мозку. Далі виникають дегенеративні процеси в м-холінергічних системах кори та гіпокампу, що супроводжуються зниженням рівня ключового ензиму синтезу ацетилхоліну. холінацетилтрансферазита характерними морфологічними змінами: дефіцитом великих пірамідних нейронів, появою скупчень білка – бере участь у міжнейронних контактах і 2) частково розщеплюючись специфічними протеазами, утворює великий N_кінцевий фрагмент, який виходить у позаклітинні середовища мозку та бере участь у процесах консолідації пам'яті. При хворобі Альтцгеймера спотворюється протеоліз білка-попередника і з середньої його частини виходить невеликий фрагмент з 41 амінокислотного залишку - власне р-амілоїд, що відкладається на поверхні нейронів. Перестає формуватися і згаданий вище великий фрагмент, що стимулює консолідацію пам'яті. Внаслідок цього виникає глибоке порушення здатності хворих до запам'ятовування.
Дегенерація великих груп нейронів, що входять до м-холінергічної системи, супроводжується також глибокими змінами низки нейропептидних систем. На жаль, поки не можна сказати, які з них первинні, а які вторинні, але очевидно дуже значне зниження рівнів кортиколіберину в потиличній корі та хвостатому ядрі, а також соматостатину у скроневій та лобовій корі. У той же час зростає рівень нейропептиду Y у так званій безіменній субстанції.
На відміну від інших психічних хвороб поки що мало інформації про лікарські засоби, що полегшують сенильную деменцію, і тому мало відповідних додаткових непрямих вказівок на біохімічні механізми хвороби. Можна лише відзначити наявні нерівноцінні дані про короткочасно полегшують хворобу засобах, що підсилюють холінергічну трансмісію, що, у разі, суперечить викладеним вище уявленням.
У порядку ілюстрації того, наскільки шкідлива поспішність у формуванні наукових гіпотез, слід згадати про те, що причиною хвороби Альтцгеймера є токсична дія алюмінію. Автори гіпотези виходили з повідомлень про високий вміст алюмінію в нейронах гіпокампу та деяких інших відділах мозку у людей, які страждали на хворобу Альтцгеймера. Цей факт був зіставлений з широким вживанням алюмінієвого посуду та алюмінієвих харчових добавок, що містять, в сучасну історичну епоху. Однак, за даними останніх робіт у цій галузі, зміни вмісту алюмінію якщо і існують, то не такі великі, як здавалося раніше, причому є підстави вважати, що проникнення в мозок та зв'язування нейронами алюмінію - це вторинний процес, що є наслідком порушення захисної функції гематоенцефалічного бар'єру-
9 . КАТІЙНИЙ БІЛОК МІЄЛІНА, ПОРУШЕННЯ ІММУНОЛОГІЧНОЇ АВТОНОМІЇ МОЗКУ, АЛЕРГІЧНИЙ ЕНЦЕФАЛОМІЄЛІТ І РОЗСІЙНИЙ СКЛЕРОЗ
Значним досягненням нейрохімії та нейроімунології є розкриття провідних механізмів низки захворювань, в основі яких лежить процес пошкодження та часткового розсмоктування мієлінової оболонки аксонів,зокрема головного мозку. Цей процес характерний для таких захворювань, як алергічний енцефаліт та розсіяний склероз, та супроводжується глибокими порушеннями функцій ЦНС, зокрема анорексією, атаксією та паралічами; завершується він значною часткою випадків летально.
Ключовим експериментом стало відкриття можливості викликати алергічний енцефаліт у різноманітних експериментальних тварин, якщо вводити їм периферично власний катіонний білок мієлінуз імуностимулятором - ад'ювантом Фрейнда.
КБМ становить близько третини білків мієліну. і являє собою великий пептид, що включає 160-190 а.о., серед яких особливо велика частка залишків основних амінокислот - лізину, аргінінуі гістидин- Близько 25% сумарно. Навіть одноразове введення цього білка в дозах менше 0,1 мг/кг з імуностимуляторами вело через 2-4 тижні до розвитку у більшості тварин алергічного енцефаліту. Було виявлено далі кілька ділянок КБМ, відповідальних за цей ефект. Мінімальними за розміром виявилися FSWGAEGQR та GSLPQKAQRPQDEN, енцефалітогенність яких близька або навіть перевищує вихідний КБМ.
В кінцевому рахунку склалося уявлення про те, що катіонний білок мієліну або його фрагменти, що вводяться тваринам системно або що надходять з мозку в загальний кровотік внаслідок якогось пошкодження гематоенцефалічного бар'єру, можуть сприйматися позамозковою імунною системою як чужорідні за умови одночасної імуностимуляції. Можливість сприйняття специфічних білків центральної нервової системи як чужорідних є наслідком часткової імунологічної автономії мозку, що виникає ще в ембріогенезі. Тому надходження катіонного білка мієліну чи його фрагментів і натомість імуностимуляції веде до розвитку проти них імунної відповіді. Зокрема відбувається специфічна активація Т-лімфоцитів; процес завершується залученням та активацією макрофагів, які й ушкоджують мієлін.
Підтримкою гіпотези про імунні механізми алергічного енцефаліту є дані про можливість його профілактики введенням великих доз тих же фрагментів катіонного білка мієліну, але без імуностимуляторів. Хоча задовільний пояснення механізму толерантності - завдання важка, очевидна специфічність явища, бо ряд пептидів і білків, близьких за змістом основних амінокислот не дають профілактичного ефекту.
Розкриття імунохімічних основ алергічного енцефаліту та близьких до нього хвороб – розсіяного склерозу та енцефаліту, що виникає іноді після вакцинації проти вірусу сказу, – змушує дослідників із зростаючою увагою ставитися до даних про імунохімічні процеси при інших психонейрологічних хворобах. Вище згадувалися припущення про можливу роль цих процесів у патогенезі епілепсії. Різноманітні спектри антитіл до мозкоспецифічних білків та ряд змін імуно-компетентних клітин реєструються при шизофренії, депресіях та ін. Очевидною є важливість накопичення та узагальнення даних, що знаходяться на стику нейрохімії та нейроімунології.
УВИСНОВКИ
1. Один з механізмів наркоманій та алкоголізму полягає в імітації наркотиком, етанолом або їх метаболітами функцій ендогенних сполук, які є в нормі факторами внутрішнього підкріплення, винагороди тощо. Вплив на їх освіту та рецепцію – перспективний шлях подолання станів абстиненції та залежності.
2. Головні ферменти метаболізму етанолу - алкогольдегідрогеназу та ацетальдегіддегідрогеназу. Модуляція їх активності впливає формування та підтримка залежності від алкоголю.
3. Основні нейрохімічні механізми хвороби Паркінсона полягають у придушенні синтезу дофаміну клітинами чорної субстанції мозку.
4. Важливими елементами нейрохімічних механізмів шизофренії є: посилення синтезу дофаміну у скроневих та деяких інших областях мозку, посилення та збочення катехоламінергічної імпульсації в напрямку від ряду структур середнього мозку до лобової кори, збочення систем видалення та окислювального розщеплення моноамінів. Фактори, що пригнічують перші два із зазначених процесів, тимчасово припиняють прояви шизофренії.
5. В основі депресивних станів лежить зниження катехоламінергічної трансмісії. Певну роль грає і збочення серотонінергічної системи. Фактори, що пригнічують зворотне захоплення та розпад катехоламінів, перешкоджають прояву депресій.
6. Важливим елементом механізмів тривожності, патологічного страху та проконфліктної поведінки може бути активація систем, що генерують та рецептують ендозепіни та похідні бета-карболіна.
7. Характерною рисою патогенезу епілепсії та ряду інших судомних станів є підвищення та збочення ефективності глутамінергічної трансмісії та порушення в ГАМК-ергічній системі. Фактори, що пригнічують першу систему і особливо активують другу, здатні усунути судомні синдроми та деякі інші прояви епілепсії.
8. Основою проявів низки старечих психозів є порушення холінергічної трансмісії. Вони пов'язані також із збоченням синтезу та патологічним накопиченням одного з білків ЦНС – бета-амілоїду.
9. Багато хвороб мозку пов'язані з імунохімічними процесами, у тому числі з порушенням імунної автономії мозку. Демієлінізуючі хвороби - алергічний енцефаліт, розсіяний склероз та ін - можуть бути викликані імунізацією до катіонного білка мієліну. При епілепсії, шизофренії та деяких інших психічних хворобах у крові виявляються антитіла до окремих нейроспецифічних білків.
Подібні документи
Сутність та наслідки розсіяного склерозу, походження його назви. Виникнення захворювання внаслідок порушення імунної системи; історія його вивчення та фактори, що сприяють появі. Найбільш поширені симптоми розсіяного склерозу.
презентація , доданий 03.12.2012
Опис розсіяного склерозу. Особливості та причини виникнення, поява бляшок у спинному та головному мозку. Спадкова схильність. Клінічні ознаки Атактичний синдром. Порушення чутливості. Типи перебігу розсіяного склерозу.
презентація , доданий 21.03.2017
Клас препаратів, виділених із макового соку для аналгезії (опіоїди). Ендогенні опіоїди, що застосовуються в періопераційному періоді, для регіонарної анестезії. Побічний ефект спинального застосування опіоїдів. Серцево-судинні ефекти опіоїдних агоністів.
контрольна робота , доданий 04.08.2009
Подання схеми нервових волокон головного та спинного мозку. Характеристика асоціативних, комісуральних (спайкових) та проекційних типів провідних шляхів. Класифікація нервових волокон за різними ознаками. Черепні та спинномозкові нерви.
презентація , доданий 27.08.2013
Загальна характеристика мозкових механізмів вищих психічних функцій, особливості системного ставлення до їх локалізації. Основні методологічні положення антилокалізаціоністів. Синдромний аналіз порушення вищих психічних функцій, його чинники.
контрольна робота , доданий 26.11.2010
реферат, доданий 21.07.2013
Особлива значимість патогенетичної терапії у клініці нервових хвороб. Типові патологічні процеси у нервовій системі. Порушення нервової трофіки. Генератори патологічно посиленого збудження. Механізм ушкодження нейронів при ішемії мозку.
лекція, доданий 13.04.2009
Вплив медикаментозних препаратів, що використовуються для лікування психічних хвороб, на вибір анестезії. Специфіка анестезіологічного посібника та ускладнення у хворих на хронічний алкоголізм, наркоманію, у стадії гострого сп'яніння, при тютюнокурінні.
реферат, доданий 13.03.2010
Спадкові та набуті порушення обміну речовин. Метаболічні енцефалопатії як розлади різних відділів центральної нервової системи. Порушення мозкового кровотоку, мієліну, нервових механізмів управління рухами та порушення рухів при пошкодженні мозочка.
реферат, доданий 13.04.2009
Шизофренія та її форми. Шизоафективний розлад. Онейроїдна кататонія. Рання дитяча шизофренія, її симптоми. Чинники ризику дитячої шизофренії. Клінічні особливості шизофренії, варіанти перебігу, характер основних розладів, можливі наслідки.
Схожі записи:
 Презентація Історія створення поеми М
Презентація Історія створення поеми М
 «Самоволка» Михайло Тірін, Сергій Лук'яненко Про книгу «Самоволка» Михайло Тірін, Сергій Лук'яненко
«Самоволка» Михайло Тірін, Сергій Лук'яненко Про книгу «Самоволка» Михайло Тірін, Сергій Лук'яненко
 План уроку на тему "навіщо ми використовуємо алфавіт" Розділ II
План уроку на тему "навіщо ми використовуємо алфавіт" Розділ II
 Девіантність, девіантна поведінка
Девіантність, девіантна поведінка
 Поняття про природний комплекс Описати невеликий природний комплекс з огляду на всі компоненти
Поняття про природний комплекс Описати невеликий природний комплекс з огляду на всі компоненти